
Mesosaurus
| Mesosaurus Temporal range:
| |
|---|---|

| |
| Holotype of Mesosaurus tenuidens (specimen MNHN 1865-77) | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | †Parareptilia |
| Order: | †Mesosauria |
| Family: | †Mesosauridae |
| Genus: | †Mesosaurus Gervais, 1865[1] |
| Species: | †M. tenuidens
|
| Binomial name | |
| †Mesosaurus tenuidens Gervais, 1865[1]
| |
| Synonyms | |
| |
Mesosaurus (meaning "middle lizard") is an extinct genus of reptile from the Early Permian of southern Africa and South America. Along with it, the genera Brazilosaurus and Stereosternum, it is a member of the family Mesosauridae and the order Mesosauria. Mesosaurus was long thought to have been one of the first marine reptiles, although new data suggests that at least those of Uruguay inhabited a hypersaline water body, rather than a typical marine environment.[3] In any case, it had many adaptations to a fully aquatic lifestyle. It is usually considered to have been anapsid, although Friedrich von Huene considered it to be a synapsid.[4] Recent study of Mesosauridae phylogeny places the group as either the basal most clade within Parareptilia or the basal most clade within Sauropsida (with the latter being the less supported position)[5] despite the skull of Mesosaurus possessing the "Synapsid condition" of one temporal fenestra.[6][7]
Discovery and naming[edit]
The holotype of M. tenuidens, MNHN 1865-77, is nicknamed the "Griqua Mesosaurus" and it was found in a Griqua hut in South Africa, likely in Kimberley, Northern Cape around 1830 and was being used as a pot lid.[1][8] The circumstances of its discovery and how it was taken from its previous owners in South Africa are unknown, but what is known is that the specimen eventually surfaced in the collection of the French palaeontologist Paul Gervais during the 1860s and he designated it as the holotype of a new genus and species he named Mesosaurus tenuidens in 1865.[1]
Since then, Mesosaurus remains have also been identified from South America and were first identified in 1908 as belonging to a second species, M. brasiliensis, by J. H. MacGregor.[9] Later studies have shown that M. brasiliensis was the same animal as M. tenuidens, which remains as the single valid species of Mesosaurus to this day.
Two other species of mesosaurids have since been described, which are Stereosternum[10] and Brazilosaurus,[11] which are also considered to be synonyms of Mesosaurus tenuidens according to Piñeiro et al. (2021).[2]
Description[edit]
Mesosaurus had a long skull that was larger than that of Stereosternum and had longer teeth. The teeth are angled outwards, especially those at the tips of the jaws.[12]
The bones of the postcranial skeleton are thick, having undergone pachyostosis. Mesosaurus is unusual among reptiles in that it possesses a cleithrum, usually found in more primitive bony fish and tetrapods. The head of the interclavicle of Mesosaurus is triangular, unlike those of other early reptiles, which are diamond-shaped.[13]
Palaeobiology[edit]
Mesosaurus was one of the first reptiles known to have returned to the water after early tetrapods came to land in the Late Devonian or later in the Paleozoic.[14] It was around 1 metre (3.3 ft) in length, with webbed feet, a streamlined body, and a long tail that may have supported a fin. It probably propelled itself through the water with its long hind legs and flexible tail. Its body was also flexible and could easily move sideways, but it had heavily thickened ribs, which would have prevented it from twisting its body.[15]
Mesosaurus had a small skull with long jaws. The nostrils were located at the top, allowing the creature to breathe with only the upper side of its head breaking the surface, in a similar manner to a modern crocodile. The teeth were originally thought to have been straining devices for the filter feeding of planktonic organisms.[15] However, this idea was based on the assumption that the teeth of Mesosaurus were numerous and close together in the jaws. Newly examined remains of Mesosaurus show that it had fewer teeth and that the dentition was suitable for catching small nektonic prey such as crustaceans.[12]
The pachyostosis seen in the bones of Mesosaurus may have enabled it to reach neutral buoyancy in the upper few meters of the water column. The additional weight may have stabilized the animal at the water's surface. Alternatively, it could have given Mesosaurus greater momentum when gliding underwater. While many features suggest a wholly aquatic lifestyle,[16] Mesosaurus may have been able to move onto land for short periods of time. Its elbows and ankles were restricted in their movement, making walking appear impossible. It is more likely that if Mesosaurus moved onto land, it would push itself forward in a similar way to living female sea turtles when nesting on beaches.[13]
Clearly amniote-type fossil embryos of Mesosaurus in an advanced stage of development (i.e. fetuses) have been discovered in Uruguay and Brazil. These fossils are the earliest record of amniote fetuses, although amniotes are inferred to have had their typical reproductive strategy since their first appearance in the Late Carboniferous. Prior to their description, the oldest known amniote fetuses were from the Triassic. One isolated coiled fetus called FC-DPV 2504 is not surrounded by calcareous eggshells, suggesting that the glands in the oviduct of Mesosaurus and probably all Paleozoic amniotes were not able to secrete calcium carbonate, in contrast to post-paleozoic archosaurs. This would explain the scarcity of egg fossils in the paleozoic amniote fossil record. One Mesosaurus specimen called MCN-PV 2214 comprises a medium-size adult with a small individual in its rib cage which is interpreted as a fetus ‘in utero’, even suggesting that Mesosaurus like many other marine reptiles, gave live birth. If this interpretation is correct, this specimen would represent the earliest known example of viviparity in the fossil record. The isolated fetus FC-DPV 2504, however, rather points to an ovoviviparous reproduction strategy in Mesosaurus.[17]
A study on vertebral column proportions suggested that, while young Mesosaurus might have been fully aquatic, adult animals spent some time on land. This is supported by the rarity of adult animals in aquatic settings, and a coprolite possessing drying fractures. However, how terrestrial these animals were is difficult to say, as their pachyostosis and other adaptations for an aquatic lifestyle would have made foraging on land difficult.[18]
Distribution[edit]
Mesosaurus was significant in providing evidence for the theory of continental drift, because its remains were found in southern Africa, Whitehill Formation, and eastern South America (Melo Formation, Uruguay and Irati Formation, Brazil), two widely separated regions.[19][20] As Mesosaurus was a coastal animal, and therefore less likely to have crossed the Atlantic Ocean, this distribution indicated that the two continents used to be joined together.
Gallery[edit]
-
 Fossil from Brazil
Fossil from Brazil -
 Fossil from Brazil
Fossil from Brazil -
 Dual fossil in Louisiana
Dual fossil in Louisiana -
 Restoration
Restoration -
![Early reconstruction of the skeleton of M. brasiliensis showing many small teeth in the jaws (MacGregor, 1908).[9]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e7/Mesosaurus.png/120px-Mesosaurus.png) Early reconstruction of the skeleton of M. brasiliensis showing many small teeth in the jaws (MacGregor, 1908).[9]
Early reconstruction of the skeleton of M. brasiliensis showing many small teeth in the jaws (MacGregor, 1908).[9] -
 Fossil of unhatched juvenile or fetus of Mesosaurus tenuidens (FC-DPV 2504) from Uruguay
Fossil of unhatched juvenile or fetus of Mesosaurus tenuidens (FC-DPV 2504) from Uruguay -
 Skeleton molds in whitish weathering shales of the Whitehill Formation, Keetmanshoop, Namibia
Skeleton molds in whitish weathering shales of the Whitehill Formation, Keetmanshoop, Namibia -
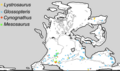 Distribution of four Permian and Triassic fossil groups used as biogeographic evidence for continental drift, and land bridging. Location of Mesosaurus remains shown by green squares
Distribution of four Permian and Triassic fossil groups used as biogeographic evidence for continental drift, and land bridging. Location of Mesosaurus remains shown by green squares




_PeerJ_4-e2036_fig_S1.png)


References[edit]
- ^ a b c d Gervais, P. (1865). Description du Mesosaurus tenuidens. Reptile fossile de l'Afrique australe. Académie des Sciences et Lettres de Montpellier. Mémoires de la Section des Sciences 6(2):169-175
- ^ a b c d Piñeiro, G.; Ferigolo, J.; Mones, A.; Núñez Demarco, P. (2021). "Mesosaur taxonomy reappraisal: are Stereosternum and Brazilosaurus valid taxa?". Revista Brasileira de Paleontologia. 24 (3): 205–235. doi:10.4072/rbp.2021.3.04. hdl:11336/175571. S2CID 244188443.
- ^ Piñeiro, G.; Ramos, A.; Goso, C.; Scarabino, F.; Laurin, M. (2012). "Unusual environmental conditions preserve a Permian mesosaur-bearing Konservat-Lagerstätte from Uruguay". Acta Palaeontologica Polonica. 57 (2): 299–318. doi:10.4202/app.2010.0113.
- ^ Huene, F. von (1940). "Osteologie und systematische Stellung von Mesosaurus". Palaeontographica Abteilung A. 92: 45–58.
- ^ Laurin, Michael; Piñeiro, Graciela (2017-11-02). "A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes". Frontiers in Earth Science. 5: 88. Bibcode:2017FrEaS...5...88L. doi:10.3389/feart.2017.00088. hdl:20.500.12008/33548.
- ^ Piñeiro, Graciela (2008). "Los mesosaurios y otros fosiles de fines del Paleozoico". In D. Perera (ed.). Fósiles de Uruguay. DIRAC, Montevideo.
- ^ Piñeiro, G.; Ferigolo, J.; Ramos, A.; Laurin, M. (2012). "Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed". Comptes Rendus Palevol. 11 (5): 379–391. Bibcode:2012CRPal..11..379P. doi:10.1016/j.crpv.2012.02.001.
- ^ Helm, Charles & Benoit, Julien. (2019). Geomythology in Southern Africa. ResearchGate 36.
- ^ a b MacGregor, J.H. (1908) Mesosaurus brasiliensis nov. sp. IN: White, I.C. (1908) Commission for Studies on Brazilian Coal Mines - Final Report; (Bilingual report, Portuguese & English), Imprensa Nacional, Rio de Janeiro, Brazil, 617 p.: Part II, pp. 301-336.
- ^ Cope, E.D. (1885). A contribution to the vertebrate paleontology of Brazil. Proceedings of the American Philosophical Society 25, 7-15.
- ^ T. Shikama and H. Ozaki (1966). "On a Reptilian Skeleton from the Palaeozoic Formation of San Paulo, Brazil". Transactions and Proceedings of the Palaeontological Society of Japan. New Series. 64: 351–358.
- ^ a b Modesto, S.P. (2006). "The cranial skeleton of the Early Permian aquatic reptile Mesosaurus tenuidens: implications for relationships and palaeobiology". Zoological Journal of the Linnean Society. 146 (3): 345–368. doi:10.1111/j.1096-3642.2006.00205.x.
- ^ a b Modesto, S.P. (2010). "The postcranial skeleton of the aquatic parareptile Mesosaurus tenuidens from the Gondwanan Permian". Journal of Vertebrate Paleontology. 30 (5): 1378–1395. Bibcode:2010JVPal..30.1378M. doi:10.1080/02724634.2010.501443. S2CID 131561248.
- ^ Laurin, Michel (2010). How Vertebrates left the Water (illustrated ed.). University of California Press. pp. xv + 199. ISBN 978-0-520-26647-6.
- ^ a b Palmer, D., ed. (1999). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. p. 65. ISBN 978-1-84028-152-1.
- ^ Canoville, Aurore; Michel Laurin (2010). "Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on paleobiological inferences". Biological Journal of the Linnean Society. 100 (2): 384–406. doi:10.1111/j.1095-8312.2010.01431.x.
- ^ Piñeiro, G.; Ferigolo, J.; Meneghel, M.; Laurin, M. (2012). "The oldest known amniotic embryos suggest viviparity in mesosaurs". Historical Biology. 24 (6): 620–630. Bibcode:2012HBio...24..620P. doi:10.1080/08912963.2012.662230. S2CID 59475679.
- ^ Pablo Nuñez Demarco et al. Was Mesosaurus a Fully Aquatic Reptile? Front. Ecol. Evol, published online July 27, 2018; doi:10.3389/fevo.2018.00109
- ^ Piñeiro, Graciela (2008). D. Perera (ed.). Fósiles de Uruguay. DIRAC, Montevideoy.
- ^ Trewick, Steve (2016). "Plate Tectonics in Biogeography". International Encyclopedia of Geography: People, the Earth, Environment and Technology. John Wiley & Sons, Ltd. pp. 1–9. doi:10.1002/9781118786352.wbieg0638. ISBN 9781118786352.
Further reading[edit]
- Parker, Steve. Dinosaurus: the complete guide to dinosaurs. Firefly Books Inc, 2003. Pg. 90
- Carroll, R. L. (1988). Vertebrate Paleontology and Evolution. W.H. Freeman and Company. ISBN 9780716718222.
- LeGrand, Homer Eugene (1988). Drifting Continents and Shifting Theories: The Modern Revolution in Geology and Scientific Change (illustrated ed.). Cambridge University Press. pp. 313. ISBN 978-0-521-31105-2.
- Margulis, Lynn; Clifford Matthews; Aaron Haselton (2000). Environmental Evolution: Effects of the Origin and Evolution of Life on Planet Earth. Contributor Clifford Matthews, Aaron Haselton (2nd ed.). MIT Press. p. 338. ISBN 978-0-262-63197-6.
- Sepkoski, Jack (2002). "A compendium of fossil marine animal genera". Bulletins of American Paleontology. 363: 5–560. ISBN 978-0-87710-450-6.






| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||