
Jebel Qatrani Formation: Difference between revisions
Adding short description: "Paleontological and geological formation in Faiyum Governorate, Egypt" |
Armin Reindl (talk | contribs) page expansion, history, geography, geology, paleoenvirontment and extensive species lists, stub tags removed Tag: Disambiguation links added |
||
| Line 1: | Line 1: | ||
{{Short description|Paleontological and geological formation in Faiyum Governorate, Egypt}} |
{{Short description|Paleontological and geological formation in Faiyum Governorate, Egypt}} |
||
{{Infobox Rockunit |
|||
{{more citations needed|date=March 2014}} |
|||
| name = Jebel Qatrani Formation |
|||
[[File:Arsinoitherium hharder.png|thumb|right|[[Heinrich Harder]]'s reconstruction of ''[[Arsinoitherium zitteli]]'' in the Oligocene grassland of [[Jebel Qatrani]] escarpment.]] |
|||
| type = [[Formation (stratigraphy)|Formation]] |
|||
The '''Jebel Qatrani Formation''' (also '''Gebel Qatrani''') is a [[paleontology|palaeontological]] and [[geologic formation]] located in the [[Faiyum Governorate]] of central [[Egypt]]. |
|||
| age = [[Eocene]]-[[Oligocene]] <br />~{{fossil range|Priabonian|Rupelian}} |
|||
| period = Eocene |
|||
| image = Jebel Qatrani Formation Stratigraphy.png |
|||
| prilithology = [[sandstone]], [[mudstone]] |
|||
| otherlithology = |
|||
| namedfor = |
|||
| namedby = |
|||
| year_ts = |
|||
| region = [[Faiyum Governorate]] |
|||
| country = {{flag|Egypt}} |
|||
| unitof = |
|||
| subunits = Lower Sequence (Lower Fossil Wood Zone) |
|||
Barite Sandstone |
|||
Conformably{{check|date=November 2012}} overlying the [[Qasr el Sagha Formation]]. It is exposed namely between the Jebel Qatrani escarpment and the Qasr el Sagha [[escarpment]], north of [[Birket Qarun]] lake near [[Faiyum]]. |
|||
Upper Sequence (Upper Fossil Wood Zone) |
|||
==Geology== |
|||
| underlies = [[Widan el Faras Basalt]] |
|||
These rocks were laid down in the [[Eocene]]-[[Oligocene]] period ([[Priabonian]] - [[Rupelian]]){{check|date=November 2012}}. This formation was originally thought to be between 35.4 and 33.3 million years old, based on initial analysis of the formation. However, analysis by Erik Seiffert in 2006 concluded that the age of the Jebel Qatrani Formation should be revised. His assessment of more recent evidence indicates an age for most of the formation of between 30.2 and 29.5 million years ago, placing it almost entirely in the [[Rupelian|Early Oligocene]]. Seiffert states that only the lowest 48 metres was laid down in the [[Eocene]],<ref name="Seiffert">{{cite journal |author=Seiffert Erik R. |title=Revised age estimates for the later Paleogene mammal faunas of Egypt and Oman|journal=Proceedings of the National Academy of Sciences of the United States of America |volume=103 |issue=13 |pages=5000–5005 |date=Jan 2006 | doi=10.1073/pnas.0600689103 |pmc=1458784 |pmid=16549773|bibcode=2006PNAS..103.5000S|doi-access=free}}</ref> but recent opinion holds the original hypothesis of a sediment straddling the Eocene-Oligocene boundary to be correct.{{Citation needed|reason=Cannot find any contradiction of Seiffert, everything found stating older appears to be from the 1990's, found one article published 2014 but that was an online republishing of Prothero&Berggren(1992)|date=October 2017}} |
|||
| overlies = [[Qasr el Sagha Formation]] |
|||
| thickness = 340 m |
|||
| extent = |
|||
| area = [[Fayum Depression]] |
|||
}} |
|||
The '''Jebel Qatrani Formation''' (also '''Gebel Qatrani''', '''Gabal Qatrani''' or '''Djebel Qatrani''') is a [[geologic formation]] located in the [[Faiyum Governorate]] of central [[Egypt]]. It is exposed between the Jebel Qatrani escarpment and the Qasr el Sagha [[escarpment]], north of [[Birket Qarun]] lake near [[Faiyum]]. The formation conformably overlies the [[Qasr el Sagha Formation]] and is topped by the [[Widan el Faras Basalt]]. The age of the formation has been subject to debate, but the most recent research indicates that it covers both the latest parts of the [[Eocene]] and the [[Early Oligocene]], spanning over the boundry between these two time periods. |
|||
The geology and fauna of this formation gives a good idea of the environment and animals present during this time period. Research suggests that the Jebel Qatrani Formation featured a mix of subtropical to [[tropical forest]], lowland [[swamp]]s and [[marsh]]es, ponds and rivers that would would empty northward into the [[Tethys Sea]]. This is supported by the presence of water-dependent fauna including [[Podocnemididae|podocnemidid turtles]], [[crocodilians]], [[sea cows]], various fish, [[Jacanidae|jacanas]], early [[Palaelodidae|flamingo-relatives]], [[ospreys]], [[herons]] and [[shoebills]]. |
|||
Mostly [[sandstone]]s deposited by meandering rivers and seasonal lakes, the lower (Eocene) part of the formation represents a woodlanded ecosystem of which much [[petrified wood]] also remains. In contrast, the upper (Oligocene) layers present indication of much drier and cooler conditions, with [[savanna]] replacing the woodland for several million years before the climate turned wetter and warmer again and the forests regrew. |
|||
Besides these, the fossil record of the Jebel Qatrani Formation is especially well known for its value to understanding the early evolution of many modern mammal groups. [[Primates]] are represented by over a dozen [[genera]], several forms of early [[Proboscidea|elephant]]s have been recovered from the sediments including the terrestrial ''[[Phiomia]]'' and the semi-aquatic ''[[Moeritherium]]''. The fossil [[rodents]] of the formation meanwhile are thought to be an important link between the African [[Phiomorpha|phiomorphs]] ([[dassie rat|dassies]], [[old world porcupine]]s, [[mole rats]] and [[cane rats]]) and the [[Caviomorpha|caviomorphs]] of [[South America]] ([[capybara]]s, [[chinchilla]]s and [[new world porcupine]]s). Besides these early members of groups that would later rise to prominence, the formation was also home to a variety of unique groups no longer found today or only found in a greatly diminished diversity. This includes the enigmatic, possibly carnivorous [[Ptolemaiida|ptolemaiids]], large [[hyaenodont]]s, a vast number of highly diverse [[hyracoids]] including species the size of rhinos, [[anthracotheres]] and the bizarre [[Embrithopoda|embrithopod]] ''[[Arsinoitherium]]''. |
|||
The [[fauna]] changes also notably during this time, seemingly coincident with the [[Eocene–Oligocene extinction event]] recorded in Eurasia. |
|||
==Geography and history== |
|||
==Fossils== |
|||
[[File:The American Museum journal (c1900-(1918)) (17971592490).jpg|left|thumb|Excavation of an ''Arsinoitherium'' skull in Fayum (1908).]] |
|||
[[File:Aegyptopithecus NT.jpg|thumb|right|Life restoration of prehistoric primate ''[[Aegyptopithecus zeuxis]]''.]] |
|||
Outcrops of the Jebel Qatrani Formation are present in the northern [[Fayum Depression]] southwest of [[Cairo]].<ref name=R87/> The Fayum Depression is an oasis west of the [[Nile]] in northern [[Egypt]].<ref name=K23>{{Cite journal|last1= Kampouridis|first1=P.|last2=Hartung|first2=J.|last3=Augustin|first3=F.J.|year=2023|title=The Eocene–Oligocene Vertebrate Assemblages of the Fayum Depression, Egypt|journal=Advances in Science, Technology & Innovation| doi=10.1007/978-3-030-95637-0_14}}</ref> |
|||
[[File:Bothriogenys fraasi.JPG|thumb|right|Reconstruction of Jebel Qatrani swamp forest at the end of the Eocene, with two ''[[Bothriogenys fraasi]]'' in front of an ''[[Arsinoitherium zitteli]]'']] |
|||
The formations of the Fayum have been studied for a significant amount of time by numerous paleontologists, with research dating back to as early as the 19th century. During this time the region was studied extensively by scientists including but not limited to [[Charles William Andrews]], [[Henry Fairfield Osborn]], [[René Fourtau]] and [[Ernst Stromer]]. Among the first names for what is now known as the Jebel Qatrani formation was the “Fluvio-Marine Series”, as coined by Hugh J. L. Beadnell. However, despite the bulk of research conducted in the late 19th and early 20th century, the Fayum localities would eventually enter a period of obscurity following the outbreak of [[World War I]] which continued throughout the mid 20th century and [[World War II]]. Research resumed during the 1960s, following an expedition under [[Elwyn LaVerne Simons]].<ref name=K23/> Previously, the formations of the region had been primarily known for the preservation of mammals, but Birds were also known from few specimens uncovered in the early 20th century. During the 60s, improved collection methods and additional expeditions by the [[Yale University|Yale]] and [[Duke University]] gathered much additional material in association with the [[Egyptian Geological Survey]] and the [[General Petroleum Company]].<ref name=R87>{{Cite journal|last1=Rasmussen|first1=D.T.|last2=Olson|first2=S.L.|last3=Simons|first3=E.L.|year=1987|title=Fossil birds from the Oligocene Jebel Qatrani formation Fayum Province, Egypt|journal=Smithsonian Contributions to Paleobiology|volume=62|pages=1-20}}</ref> Around the 70s, the scope of the expedition was broadened to account for more diverse fields of study, leading to more precise datings of the strata.<ref name=K23/> |
|||
===Mammals=== |
|||
* ''[[Arsinoitherium zitteli]]'', an arsinoitheriid |
|||
==Geology and stratigraphy== |
|||
* ''[[Bothriogenys fraasi]]'', an anthracotheriid |
|||
[[File:Geological section Gebel Qatrani formation.png|thumb|Geological section Gebel Qatrani formation]] |
|||
* ''Herodotius pattersoni'', a [[pseudoungulate]] |
|||
The formation overlies the Eocene [[Qasr el Sagha Formation]] and is overlaid by the Oligocene [[Widan el Faras Basalt]]. The formation contains at least two major fossil bearing layers, one in the upper sequence, which is used to refer to the top layers of the formation, and a second in the lower sequence. These two layers have also been called the Upper and Lower Fossil Wood Zones in older publications. Both sequences of the Jebel Qatrani Formation are separated from one another by the so called Barite Sandstone, a layer with a thickness of {{convert|4-10|m|abbr=on}}.<ref name=R87/><ref name=K23/> |
|||
* ''[[Plesiopithecus]]'', an enigmatic lemur-like primate |
|||
* [[Pliohyracidae]], large [[hyrax]]es |
|||
The dating of the Jebel Qatrani Formation has historically been under debate, with some researchs having placed it either entirely within the Eocene or Oligocene and some arguing that it spans both periods. Rassmussen and colleagues for instance argued that the central Barite Sandstones separating both fossil bearing layers mark the exact [[Eocene-Oligocene Boundry]]. Part of the reasoning for this connects to the primate fauna of the formation, with propliopithecids and parapithecines only appearing in the upper localities. Other dating methods meanwhile have generally not been applicable. According to Seiffert, the mammal fauna of the formation on its own is too endemic and marine invertebrates are absent, preventing biostratigraphic dating. Radioisotopic dating was conducted on the overlying Widan el Faras Basalt, however the results of this suggested an age of 23.6 million years for its lower units, much younger than prior estimates for the Jebel Qatrani Formation. However, in a 2006 publication Seiffert draws comparisson between the Fayum fauna and the fossil record of the [[Ashawq Formation]] in [[Oman]], which notably preserved vertebrates as well as [[foraminifera]] useful in dating. Foraminifera biostrategraphy and magnetostratigraphy suggest that the examined localities in Oman date to approximately 31-31.5 (Taqah locality) and 33.7-33.3 Ma (Thaytiniti locality). Seiffert argues that the mammal fauna from Oman most closely resembles those found in the oldest localities of the Lower Sequence of the Jebel Qatrani Formation (L-41) and the oldest localities of the Upper Sequence (quarries G and V). The hyraxes ''[[Thyrohyrax]]'' and ''[[Saghatherium]]'' occur in both formations, as does the primate ''[[Moeripithecus]]''. More generally, both formations preserve [[Propliopithecidae|propliopithecids]] and [[Parapithecinae|parapithecine]] [[Parapithecidae|parapithecids]] as well as [[Oligopithecidae|oligopithecids]]. While the former two groups are restricted to the younger Fayum sequence, the latter is the most common primate family in the older sediments. Assuming the traditional interpretation of the Jebel Qatrani Formation, this would mean that many of these taxa would have had to appear 2 to 4 million years earlier in Egypt than in Oman, which is considered to be unlikely by Seiffert. According to them the formation spans approximately 8 million years, with its oldest localities situated in latest Eocene strata. Locality BQ-2 has been estimated to be 37 million years old (early [[Priabonian]]), while L-41 falls into an age range of 34.8–33.7 million years old, a timespan that includes the Eocene-Oligocene Boundry. Although an earliest Oligocene age could not be disproven by Seiffert, he argues that a latest Eocene age should be considered more likely based on an unconformity present just above the locality (one also acknowledged as a possible candidate for the EOB by Rassmussen). This means that only the lower {{convert|48|m|abbr=on}} of the formation are Eocene in age, including both the BQ-2 and L-41 localities. The remainder of the Lower Sequence, as well as the entire Upper Sequence, would subsequently fall within the Oligocene. The position of quarries A and B in regards to the boundry is ambiguous, however Quarry E on the other hand is considered without doubt Oligocene (ca. 33 Ma) in age by Seiffert.<ref name="Seiffert">{{cite journal |author=Seiffert Erik R. |title=Revised age estimates for the later Paleogene mammal faunas of Egypt and Oman|journal=Proceedings of the National Academy of Sciences of the United States of America |volume=103 |issue=13 |pages=5000–5005 |date=Jan 2006 | doi=10.1073/pnas.0600689103 |pmc=1458784 |pmid=16549773|bibcode=2006PNAS..103.5000S|doi-access=free}}</ref> |
|||
* [[Ptolemaiida]], carnivorous [[Afrotheria|afrothere]] mammals |
|||
* ''[[Qatranilestes]]'' and ''[[Widanelfarasia]]'', relatives of [[golden mole]]s, [[otter shrew]]s, and [[tenrec]]s |
|||
==Paleoenvironment== |
|||
* Several [[hyaenodontid]]s (''[[Akhnatenavus]] cf. leptognathus, [[Metapterodon]]'') |
|||
[[File:Arsinoitherium hharder.png|thumb|left|Arsinoitherium illustrated by Heinrich Harder.]] |
|||
* Several [[hystricognath]] rodents (e.g. ''[[Acritophiomys]] bowni, A. woodi, [[Elwynomys]] lavocati, [[Gaudeamus (rodent)|Gaudeamus]] hylaeus'') |
|||
The environment of the Jebel Qatrani formation has been described as a [[subtropical]] to [[tropical]] lowland plain by Bown, who further suggests the presence of streams and ponds. Based on the fossil bird remains, which includes the fossils of a variety of animals highly associated with water (ospreys, early flamingos, jacanas, herons, storks, cormorants and shoebills), Rasmussen and colleages inferred that the environment featured slow-moving freshwater with a substantial amount of aquatic vegetation, which matches the prior hypothesis. Although [[lithology]] suggests that most fossils were deposited on sandbanks after being transported by currents, the authors argue that [[swamps]] could have easily formed along the banks of the river that was present during the Oligocene and may account for the mudstone found in certain quarries. They furthermore suggest that the fossil birds of Fayum, due to their affinities with modern groups, should be considered a more valuable indicator of the environment when compared with the fossil mammals, many of which belonged to families lacking modern examples. The absence of other birds typical for such an environment may be explained either through sampling bias or due to the fact that said groups had simply not yet been present in Oligocene Africa. Generally, Rasmussen and colleagues compare the environment of Jebel Qatrani to freshwater habitats in modern Central Africa.<ref name=R87/> The discovery of snakehead fossils seem to support Rasmussen's interpretation, as the genus ''Parachanna'' today prefers slow-moving backwaters with plenty of vegetation. Other fish present meanwhile, notably ''Tylochromis'', suggest that deep, open water was likewise present. The river channels may have been overgrown with [[reed (plant)|reeds]], [[Cyperus papyrus|papyrus]] and featured floating vegetation like [[Nymphaeaceae|water lilies]] and ''[[Salvinia]]''.<ref name=M04/> In a 2001 paper Rasmussen ''et al.'' argued that the sandstone and mudstone of the formation likely formed as sediments were [[Aggradation|aggraded]] by a system of river channels that emptied towards the west into the Tethys. Here they reconstructed the environment as a tropical lowland swamp forest intermingled with marshes. They furthermore suggest that the environment would have experienced [[monsoon]]s.<ref name=R01/> Overall this indicates that this region was a part of an extensive belt of tropical forest that stretched across what is now northern Africa, which would gradually give rise to open woodland and even steppe the further one was to travel inland.<ref name=K23/> |
|||
* A primitive [[marsupial]], maybe related to ''[[Peratherium]]'' |
|||
* Primitive Old World [[monkey]]s (e.g. ''[[Aegyptopithecus zeuxis]], [[Biretia]] fayumensis'' & ''B. megalopsis'') |
|||
==Paleobiota== |
|||
===Fish=== |
|||
'''Actinopterygii''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Alestidae]] indet.<ref name=M04>{{Cite journal|last1=Murray|first1=A.M.|year=2004|title=Late Eocene and early Oligocene teleost and associated ichthyofauna of the Jebel Qatrani Formation, Fayum, Egypt.|url=https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.0031-0239.2004.00384.x|journal=Palaeontology|volume=47|issue=3|pages=711-724|doi=10.1111/j.0031-0239.2004.00384.x}}</ref> |
|||
| |
|||
|Lower and Upper Sequence |
|||
|Quarry E, I, M, R & X |
|||
|Various teeth |
|||
|Different teeth belonging to [[Characiformes]], they are common across both fossil bearing members. A large range of tooth morphotypes is present in the formation, however this may not reflect the actual diversity of african tetra species present. |
|||
| |
|||
|- |
|||
|[[Amiiformes]] indet.<ref name=M04/> |
|||
| |
|||
| |
|||
| |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|''[[Lates]]''<ref name=M04/> |
|||
|''Lates sp.'' |
|||
|Lower and Upper Sequence |
|||
|Quarry E, I, M & V |
|||
|Two crania and various other isolated material. |
|||
|Its absence from the lacustrine L-41 environment might indicate that the water was too shallow. Modern ''Lates'' can grow large and prefer large lakes and rivers. |
|||
| |
|||
|- |
|||
|''[[Parachanna]]''<ref name=M04/> |
|||
|''[[Parachanna fayumensis]]'' |
|||
|Lower and Upper Sequence |
|||
|Quarry L-14, P & M |
|||
|Various cranial remains including a dentary. |
|||
|A snakehead fish, the Jebel Qatrani material may suggest that this group arrived in Africa earlier than it had been previously thought. |
|||
| |
|||
|- |
|||
|[[Siluriformes]] indet.<ref name=M04/> |
|||
| |
|||
|Lower and Upper Sequence |
|||
|Quarry L-41, B, G, E, I, J, K, M, P & V |
|||
|Three skulls and various postcranial remains, predominantly fin spines. |
|||
|The three known skulls are approximately the same size, but may not represent the same taxon. The postcranial elements indicate a greater size range. In some aspects the Jebel Qatrani catfish resemble the genus ''[[Fajumia]]''. |
|||
| |
|||
|- |
|||
|cf. ''[[Tylochromis]]''<ref name=M04/> |
|||
| |
|||
|Lower and Upper Sequence |
|||
|Quarry E, I & X |
|||
|A lower jaw and additional material including teeth. |
|||
|A basal [[Cichlidae|cichlid]]. |
|||
| |
|||
|- |
|||
|} |
|||
'''Chondrichthyes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Dasyatoidea]] indet.<ref name=M04/> |
|||
| |
|||
| |
|||
| |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|[[Lamniformes]] indet.<ref name=M04/> |
|||
| |
|||
| |
|||
| |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|} |
|||
'''Sarcopterygii''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Dipnoi]] indet.<ref name=M04/> |
|||
| |
|||
| |
|||
|Quarry L-41 |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|} |
|||
===Reptiles=== |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Albertwoodemys]]''<ref name=GMW11>{{Cite journal |last1=Gaffney|first1=E.S.|last2=Meylan|first2=P.A.|last3=Wood|first3=R.C.|last4=Simons|first4=E.|last5=de Almeida Campos|first5=D.|year=2011 |title=Evolution of the side-necked turtles: the family Podocnemididae |journal=Bulletin of the American Museum of Natural History |volume=350 |pages=1–237 |hdl=2246/6110| doi=10.1206/350.1| s2cid=83775718 }}</ref> |
|||
|''Albertwoodemys testudinum'' |
|||
|Lower Sequence |
|||
|A locality west of the AMNH quarries. |
|||
|A partial plastron with articulated peripheral elements of the upper shell. |
|||
|A side-necked turtle of the family [[Podocnemididae]] with a high-domed shell likely similar to tortoises. It may be a related taxon to specimen UCMP 42008, from the Miocene of Kenya. |
|||
| |
|||
|- |
|||
|''[[Andrewsemys]]''<ref name=GMW11/><ref name=PG17>{{Cite journal|last1=Pérez-García|first1=A.|year=2017|title=New information and establishment of a new genus for the Egyptian Paleogene turtle “Stereogenys” libyca (Podocnemididae, Erymnochelyinae)|journal=Historical Biology|pages=1–10|doi=10.1080/08912963.2017.1374383}}</ref> |
|||
|''Andrewsemys libyca'' |
|||
| |
|||
|North of [[Lake Qarun]] |
|||
|Various shell remains. |
|||
|A side-necked turtle of the family Podocnemididae previously known as ''"Stereogenys" lybica''. The holotype specimen, among the best preserved turtle shells from Fayum, was thought to be lost before being rediscovered and used to errect a new genus. Several additional specimens are known from the Jebel Qatrani and its underlying formations. |
|||
| |
|||
|- |
|||
| |
|||
|''[[Crocodylus megarhinus|"Crocodylus" megarhinus]]''<ref name=BG20>{{Cite journal|last1=Brochu|first1=C.A.|last2=Gingerich|first2=P.D.|year=2000|title=New tomistomine crocodylian from the middle Eocene (Bartonian) of Wadi Hitan, Fayum Province, Egypt|url=https://deepblue.lib.umich.edu/bitstream/handle/2027.42/48660/ID527.pdf?sequence=2|journal=CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN|volume=30|issue=10|pages=251-268}}</ref> |
|||
| |
|||
| |
|||
|Various cranial and mandibular remains |
|||
|A large and broad-snouted species of crocodilian incorrectly assigned to ''Crocodylus''. A second Fayum crocodile, ''"Crocodylus" articeps'', has been considered a younger individual of this species by Christopher Brochu. |
|||
| |
|||
|- |
|||
|''[[Dacquemys]]''<ref name=GMW11/><ref name=E02>{{Cite journal|last1=Gaffney|first1=E.S.|last2=Deblieux|first2=D.D.|last3=Simons|first3=E.L.|last4=Sánchez-Villagra|first4=M.R.|last5=Meylan|first5=P.A.|year=2002|title=Redescription of the Skull of Dacquemys, a Podocnemidid Side-Necked Turtle from the Late Eocene of Egypt|url=https://www.researchgate.net/profile/Donald-Deblieux/publication/232687745_Redescription_of_the_Skull_of_Dacquemys_Williams_1954_a_Podocnemidid_Side-Necked_Turtle_from_the_Late_Eocene_of_Egypt/links/56378af808aed65d3c42c794/Redescription-of-the-Skull-of-Dacquemys-Williams-1954-a-Podocnemidid-Side-Necked-Turtle-from-the-Late-Eocene-of-Egypt.pdf|journal=American Museum Novitates|pages=1-16|doi=10.1206/0003-0082(2002)372<0001:ROTSOD>2.0.CO;2}}</ref> |
|||
|''Dacquemys paleomorpha'' |
|||
|Lower Sequence |
|||
|Quarry B |
|||
|A nearly complete skull |
|||
|A side-necked turtle of the family [[Podocnemididae]] that may be a related taxon to specimen UCMP 42008. It is possible that ''Dacquemys'' represents skull material of ''Albertwoodemys''. |
|||
| |
|||
|- |
|||
|''[[Eogavialis]]''<ref name=BG20/> |
|||
|''Eogavialis africanum'' |
|||
| |
|||
| |
|||
| |
|||
|An early genus of [[Gavialoidea|gavialoid]] common in the formation. Two species have originally been named from Jebel Qatrani,''Eogavialis gavialoides'' and ''Eogavialis tenuirostre'' (both originally as ''[[Tomistoma]]''), however recent research suggests that all Fayum ''Eogavialis'' species including those of other formations in the region may simply represent a single species.<ref name=K23/> |
|||
| |
|||
|- |
|||
|''[[Gigantochersina]]''<ref name=HP03>{{Cite journal|last1=Holroyd|first1=P.A.|last2=Parham|first2=J.F.|year=2003|title=The antiquity of African tortoises.|journal=Journal of Vertebrate Paleontology| volume=23|issue=3|pages=688-690|doi=10.1671/1870}}</ref> |
|||
|''Gigantochersina ammon'' |
|||
|Lower Sequence |
|||
|Quarry A & B |
|||
|Multiple specimens including a nearly complete carapace and plastron as well as a partial pelvis |
|||
|The oldest known tortoise from Africa. ''Testudo beadnelli'' and ''Testudo isis'' were both named, but have since then been synonymized with ''G. ammon''.<ref name=K23/> |
|||
| |
|||
|- |
|||
|''[[Neochelys]]''<ref name=GMW11/> |
|||
|''Neochelys fajumensis'' |
|||
|Lower and Upper Sequence |
|||
|Quarry L-41, A, B, C, I, M, O, P & R |
|||
|Various remains of the plastron and carapace as well as skull material. |
|||
|A side-necked turtle of the family Podocnemididae, similar to ''[[Erymnochelys]]'' and previously assigned to said genus. The type specimen, an anterior plastron, has potentially been lost. They are among the most common fossils in the quarries L-41, I and M. |
|||
| |
|||
|- |
|||
|[[Varanidae]] indet.<ref name=K23/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|A presacral vertebra |
|||
|I single bone described by Smith ''et al.'' and originally assigned to the genus ''Varanus''. Later research by Holmes ''et al.'' suggests it was a stem-varanid, distinct from other remains found in the formation. |
|||
| |
|||
|- |
|||
|''[[Varanus]]''<ref name=K23/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry I & M |
|||
|Vertebrae |
|||
|Isolated bones described by Holmes ''et al.'' and considered to be part of the modern genus ''Varanus''. They argue that this represents the oldest known member of the genus and indicates an African origin for modern monitor lizards. |
|||
| |
|||
|- |
|||
|} |
|||
===Birds=== |
===Birds=== |
||
'''Accipitriformes''' |
|||
* ''[[Eremopezus]]'', an enigmatic large groundbird |
|||
{| class="wikitable" align="center" width="100%" |
|||
* ''[[Goliathia]]'', a large shoebill relative |
|||
|- |
|||
* ''[[Palaeoephippiorhynchus]] dietrichi'', a large stork |
|||
! Name |
|||
* ''Xenerodiops'', an unusual [[heron]] |
|||
! Species |
|||
* A [[cormorant]]-like bird, possibly related to genus ''Piscator'' |
|||
! Member |
|||
* Several [[Jacanidae|jacana]]s |
|||
! Locality |
|||
* One or two species of primitive [[osprey]] and another [[bird of prey]] (possibly an ancestral [[sea eagle]]) |
|||
! Material |
|||
* A [[turaco]] |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Accipitridae]] indet., aff. ''[[Haliaeetus]]''<ref name=R87/> |
|||
| |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|The distal end of a tarsometatarsus. |
|||
|A bird of prey similar in size and morphology to the modern sea eagles. The early strata of the formation indicate closer proximity to the shore, possibly supporting a lifestyle similar to modern ''Haliaeetus'' species. |
|||
| |
|||
|- |
|||
|[[Pandionidae]] indet., aff. ''[[Pandion (genus)|Pandion]]''<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|The distal end of a humerus. |
|||
|A fossil nearly identical to the extant [[osprey]], but smaller. |
|||
| |
|||
|- |
|||
|Pandionidae? indet.<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|A damaged carpometacarpus. |
|||
|A raptorial bird approximately the size of the extant osprey, however more robust. |
|||
| |
|||
|- |
|||
|} |
|||
'''Charadriiformes''' |
|||
===Other animals=== |
|||
{| class="wikitable" align="center" width="100%" |
|||
* ''[[Albertwoodemys]]'', a side-necked turtle |
|||
|- |
|||
* ''[[Crocodylus megarhinus|"Crocodylus" megarhinus]]'', an ancestral crocodile |
|||
! Name |
|||
* ''[[Eogavialis africanum]]'', a primitive gharial |
|||
! Species |
|||
* ''[[Parachanna fayumensis]]'', a snakehead fish |
|||
! Member |
|||
* ''Pterosphenus'', a [[snake]] |
|||
! Locality |
|||
* [[African tetras]] (Alestidae) or similar [[Characiformes]] |
|||
! Material |
|||
* [[Catfish]], perhaps of genus ''Fajumia'' |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Janipes]]''<ref name=R87/> |
|||
|''Janipes nymphaeobates'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|The distal end of a tarsometatarsus |
|||
|A large species of [[Jacanidae|jacana]], exceeding all modern taxa in size but still smaller than ''Nupharanassa bulotorum''. The Fayum jacanas already show signs of a lifestyle like that of their modern relatives and subsequently may indicate a dense floating vegetation. |
|||
| |
|||
|- |
|||
| rowspan=2 | |
|||
''[[Nupharanassa]]''<ref name=R87/> |
|||
|''Nupharanassa bulotorum'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|Multiple specimens preserving the tarsometatarsus. |
|||
|A large species of jacana, between 30-35% larger than the largest extant species, the [[bronze-winged jacana]]. |
|||
| |
|||
|- |
|||
|''Nupharanassa tolutaria'' |
|||
|Lower Sequence |
|||
|Quarry E |
|||
|The distal end of a tarsometatarsus. |
|||
|A smaller relative of ''Nupharanassa bulotorum'', it's distinguished by its much smaller size and older age. It is the only known jacana in the lower sequence of the Jebel Qatrani Formation. |
|||
| |
|||
|- |
|||
|} |
|||
'''Ciconiiformes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Palaeoephippiorhynchus]]''<ref name=R87/> |
|||
|''Palaeoephippiorhynchus dietrichi'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A nearly complete rostrum with mandible and partial cranium and the distal end of a right tibiotarsus. |
|||
|A large stork comparable in size to the modern [[jabiru]] and [[marabou stork]]. While the type material bears great resemblance to the extant [[saddlebill]], a tibiotarsus found in a different quarry shares no similarities with any modern taxa and may or may not belong to ''Palaeoephippiorhynchus''. |
|||
| |
|||
|- |
|||
|} |
|||
'''Cuculiformes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Musophagidae]] indet., aff. ''[[Crinifer]]''<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|The distal ends of a tarsometatarsus and a humerus. |
|||
|A relatively large type of [[turaco]] and among the earliest recorded members of the group. It shares similarities to the extant genus ''Crinifer''. |
|||
| |
|||
|- |
|||
|} |
|||
'''Gruiformes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Gruidae]] indet.<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|The distal end and partial shaft of a tarsometatarsus. |
|||
|A bird bearing resemblance to modern [[crowned crane]]s, cranes of the genus ''[[Grus (genus)|Grus]]'' and the [[Aramus (bird)|limpkin]]. It's size is between that of the limpkin and the [[demoiselle crane]], although closer to the former. |
|||
| |
|||
|- |
|||
|[[Rallidae]] indet.<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|The distal end of a tarsometatarsus. |
|||
|Possibly a type of rail bearing resemblance to the genera ''[[Sarothrura]]'', ''[[Coturnicops]]'' and ''[[Laterallus]]''. |
|||
| |
|||
|- |
|||
|} |
|||
'''Paleognathae''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Eremopezus]]''<ref name=R87/><ref name=R01>{{Cite journal|last1=Rasmussen|first1=D. T.|last2=Simons|first2=E.L.|last3=Hertel|first3=F.|last4=Judd|first4=A.|year=2001|title=Hindlimb of a giant terrestrial bird from the upper Eocene, Fayum, Egypt.|url=https://onlinelibrary.wiley.com/doi/pdf/10.1111/1475-4983.00182|journal=Palaeontology|volume=44|issue=2|pages=325-337|doi=10.1111/1475-4983.00182}}</ref> |
|||
|''Eremopezus eocaenus'' |
|||
|Lower Sequence |
|||
|Various fragmentary leg bones including the distal end of a tibiotarsus and the distal end of a tarsometatarsus. |
|||
|An enigmatic large groundbird comparable in size to a modern [[rhea (bird)|rhea]]. It was initially thought to be a type of [[ratite]], but its relationship with other birds has repeatedly been questioned. ''Stromeria fajumensis'', found in the same formation, is thought to be synonymous with ''Eremopezus''. |
|||
| |
|||
|- |
|||
|} |
|||
'''Pelecaniformes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Ardeidae]] indet.<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry I & M |
|||
|A partial rostrum, a distal tarsometatarsus of a juvenile specimen and the first phalanx of the third toe. |
|||
|A medium sized heron comparable in size to the [[reddish egret]] and the [[great white egret]]. The lack of overlapping material means that it is uncertain if these fossils represent one or several species. |
|||
| |
|||
|- |
|||
|''[[Goliathia]]''<ref name=R87/> |
|||
|''Goliathia andrewsi'' |
|||
|Lower and Upper Sequence |
|||
|Quarry M |
|||
|A complete ulna and the distal end of a tarsometatarsus. |
|||
|A large relative of the modern [[shoebill]]. |
|||
|[[File:Goliathia andrewsii HD.jpg|frameless|center]] |
|||
|- |
|||
|''[[Nycticorax]]''<ref name=R87/> |
|||
|Nycticorax sp. |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A nearly complete tarsometatarsus and a coracoid. |
|||
|A bird who's remains are described as being identical to that of the extant [[black-crowned night heron]], indicating that night herons diverged from other herons at least 31 million years ago. |
|||
| |
|||
|- |
|||
|''[[Xenerodiops]]''<ref name=R87/> |
|||
|''Xenerodiops mycter'' |
|||
|Upper Sequence |
|||
|Quarry I & M |
|||
|A nearly complete rostrum and a humerus lacking its distal end. |
|||
|An unusual heron with a heavy and downwards curved beak. |
|||
| |
|||
|- |
|||
|} |
|||
'''Phoenicopteriformes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Phoenicopteridae]] indet. Species 1, aff. ''[[Palaelodus]]''<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M & I |
|||
|Multiple specimens including the tibiotarsus and parts of the metacarpals. |
|||
|Several bones of the Fayum phoenicopteriform show both similarities to derived flamingos as well as to the more basal ''Palaeolodus'', which Rasmussen and colleagues considered Phoenicopterids rather than placing it within its own family. The material suggests a bird approximately the size of ''Palaelodus gracilipes''. |
|||
| |
|||
|- |
|||
|Phoenicopteridae indet. Species 2<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A coracoid fragment |
|||
|A second, larger phoenicopteriform, closer in size to ''Palaeolodus crassipes'' and the [[lesser flamingo]]. |
|||
| |
|||
|- |
|||
|} |
|||
'''Suliformes''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Phalacrocoracidae]] indet.<ref name=R87/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A distal premaxilla. |
|||
|An indetermined cormorant showing a more gradually tapering and more strongly hooked beak. Among modern cormorants, it most closely resembles the [[Guanay cormorant]]. |
|||
| |
|||
|- |
|||
|} |
|||
===Mammals=== |
|||
'''Afroinsectivora''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Eochrysochloris]]''<ref name=S10>{{Cite journal|last1=Seiffert|first1=E.R.|year=2010|title=The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt.|url=https://bioone.org/journals/acta-palaeontologica-polonica/volume-55/issue-4/app.2010.0023/The-Oldest-and-Youngest-Records-of-Afrosoricid-Placentals-from-the/10.4202/app.2010.0023.full|journal= Acta Palaeontologica Polonica|volume=55|issue=4|pages=599-616|doi=10.4202/app.2010.0023}}</ref> |
|||
|''Eochrysochloris tribosphenus'' |
|||
|Lower Sequence |
|||
|Quarry E |
|||
| |
|||
|A tiny member of the [[Afrosoricida]]. |
|||
| |
|||
|- |
|||
|''[[Herodotius]]''<ref name)SHB91>{{Cite journal|last1=Simons|first1=E.L.|last2=Holroyd|first2=P.A.|last3=Bown|first3=T.M.|year=1991|title=Early tertiary elephant-shrews from Egypt and the origin of the Macroscelidea.|url=https://www.pnas.org/doi/pdf/10.1073/pnas.88.21.9734|journal=Proceedings of the National Academy of Sciences|volume=88|issue=21|pages=9734-9737}}</ref> |
|||
|''Herodotius pattersoni'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Fossils of the dentary and maxilla. |
|||
|A species of [[elephant shrew]]. |
|||
| |
|||
|- |
|||
|''[[Jawharia]]''<ref name=P19/><ref name=S10/> |
|||
|''Jawharia tenrecoides'' |
|||
|Lower Sequence |
|||
|Quarry E |
|||
| |
|||
|A relative of [[tenrec]]s and [[golden mole]]s. |
|||
| |
|||
|- |
|||
|''[[Metoldobotes]]''<ref name=K23/> |
|||
|''Metoldobotes stromeri'' |
|||
| |
|||
| |
|||
| |
|||
|A genus of elephant shrew. |
|||
| |
|||
|- |
|||
|''[[Qatranilestes]]''<ref name=P19>{{Cite journal|last1=Pickford|first1=M.|year=2019|title=Tiny Tenrecomorpha (Mammalia) from the Eocene of Black Crow, Namibia.|url=https://mme.gov.na/files/publications/121_GSN%20Comms%2021_2_Pickford%20Nanogale.pdf|journal=Communications of the Geological Survey of Namibia|volume=21|pages=15-25}}</ref><ref name=S10/> |
|||
|''Qatranilestes oligocaenus'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|A badly preserved mandible. |
|||
|The most derived member of the Afrosoricida in the formation. |
|||
| |
|||
|- |
|||
|rowspan=2| |
|||
''[[Widanelfarasia]]''<ref name=P19/><ref name=S10/> |
|||
|''Widanelfarasia bowni'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|A stem-afrosoricid. |
|||
| |
|||
|- |
|||
|''Widanelfarasia rasmusseni'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|} |
|||
'''Artiodactyla''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|rowspan=4| |
|||
''[[Bothriogenys]]''<ref name=S15>{{Cite journal|last1=Sileem|first1=A.H.|last2=Sallam|first2=H.M.|last3=Hewaidy|first3=A.G.A.|last4=Gunnell|first4=G.F.|last5=Miller|first5=E.R.|year=2015|title= Anthracotheres (Mammalia, Artiodactyla) from the upper-most horizon of the Jebel Qatrani formation, latest Early Oligocene, Fayum depression, Egypt|url= https://www.researchgate.net/profile/Afifi-Sileem/publication/296700340_ANTHRACOTHERES_MAMMALIA_ARTIODACTYLA_FROM_THE_UPPER-MOST_HORIZON_OF_THE_JEBEL_QATRANI_FORMATION_LATEST_EARLY_OLIGOCENE_FAYUM_DEPRESSION_EGYPT/links/56e5e56d08ae65dd4cc0da28/ANTHRACOTHERES-MAMMALIA-ARTIODACTYLA-FROM-THE-UPPER-MOST-HORIZON-OF-THE-JEBEL-QATRANI-FORMATION-LATEST-EARLY-OLIGOCENE-FAYUM-DEPRESSION-EGYPT.pdf|journal=Egyptian Journal of Paleontology|volume=15| pages=1-11|issn=1687 - 4986}}</ref><ref name=H96/><ref name=SG22>{{Cite journal|last1=Sileem|first1=A.H.|last2=Abu El‐Kheir|first2=G.A.|year=2022|title=Complete skull of Bothriogenys fraasi (Mammalia, Artiodactyla, Anthracotheriidae) from the Early Oligocene, Fayum, Egypt.|url=https://www.researchgate.net/profile/Gebely-Abu-El-Kheir-3/publication/363479763_Complete_skull_of_Bothriogenys_fraasi_Mammalia_Artiodactyla_Anthracotheriidae_from_the_Early_Oligocene_Fayum_Egypt/links/631ed7dc071ea12e36294c20/Complete-skull-of-Bothriogenys-fraasi-Mammalia-Artiodactyla-Anthracotheriidae-from-the-Early-Oligocene-Fayum-Egypt.pdf|journal=Geological Journal|doi=10.1002/gj.4574}}</ref><ref name=OLPC12>{{Cite journal|last1=O'Leary|first1=M.A.|last2=Patel|first2=B.A.|last3=Coleman|first3=M.N.|year=2012|title=Endocranial petrosal anatomy of Bothriogenys (Mammalia, Artiodactyla, Anthracotheriidae), and petrosal volume and density comparisons among aquatic and terrestrial artiodactyls and outgroups|url=https://www.researchgate.net/profile/Biren-Patel-2/publication/261971393_Endocranial_Petrosal_Anatomy_of_Bothriogenys_Mammalia_Artiodactyla_Anthracotheriidae_and_Petrosal_Volume_and_Density_Comparisons_Among_Aquatic_and_Terrestrial_Artiodactyls_and_Outgroups/links/542f34920cf277d58e91ef26/Endocranial-Petrosal-Anatomy-of-Bothriogenys-Mammalia-Artiodactyla-Anthracotheriidae-and-Petrosal-Volume-and-Density-Comparisons-Among-Aquatic-and-Terrestrial-Artiodactyls-and-Outgroups.pdf|journal=Journal of Paleontology|volume=86|issue=1|pages=44-50|doi=10.1666/10-091.1}}</ref> |
|||
|''Bothriogenys andrewsi'' |
|||
|Upper Sequence |
|||
|L-75 |
|||
|Teeth and dentary remains. |
|||
|A large [[Anthracotheriidae|anthracothere]] from the uppermost levels of the formation. |
|||
|rowspan=4| |
|||
[[File:Bothriogenys fraasi.JPG|frameless|center|''Bothriogenys fraasi'' (foreground)]] |
|||
|- |
|||
|''Bothriogenys fraasi'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
| |
|||
|A species of ''Bothriogenys'' arising from ''B. gorringei''. It may have either split into two lineages or given rise to ''B. andrewsi'' through [[anagenesis]]. |
|||
|- |
|||
|''Bothriogenys gorringei'' |
|||
|Lower Sequence |
|||
|Quarry M |
|||
|Originally described based on a mandible. |
|||
|The oldest species of ''Bothriogenys'' in Jebel Qatrani, it may have split into two lineages giving rise to ''B. fraasi'' and ''B. rugulosus''. It may have been a semi-aquatic browser or grazer, feeding primarily on foliage. |
|||
|- |
|||
|''Bothriogenys rugulosus'' |
|||
|Upper Sequence |
|||
| |
|||
| |
|||
|A species of ''Bothriogenys'' that likely evolved from ''B. gorringei''. |
|||
|- |
|||
|''[[Nabotherium]]''<ref name=SG22/><ref name=SSHMG16>{{Cite journal|last1=Sileem|first1=A.H.|last2=Sallam|first2=H.M.|last3=Hewaidy|first3=A.G.A.|last4=Miller|first4=E.R.|last5=Gunnell|first5=G.F.| year=2016|title=A new anthracothere (Artiodactyla) from the early Oligocene, Fayum, Egypt, and the mystery of African ‘Rhagatherium’ solved|journal=Journal of Paleontology|volume=90|issue=1|pages= 170–181|doi=10.1017/jpa.2016.13}}</ref><ref name=K23/> |
|||
|''Nabotherium aegyptiacum'' |
|||
| |
|||
| |
|||
|A partial skull, several fossils of the mandible and maxilla as well as isolated teeth. |
|||
|An anthracothere with well developed canine teeth. Its dentition is better suited to crushing fruit rather than slicing vegetation. |
|||
| |
|||
|- |
|||
|''[[Qatraniodon]]''<ref name=S15/><ref name=SG22/><ref name=GM>Patricia A. Holroyd, Fabrice Lihoreau, Gregg F. Gunnell und Ellen R. Miller: Anthracotheriidae. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 843–851</ref> |
|||
|''Qatraniodon parvus'' |
|||
|Lower Sequence |
|||
| |
|||
|The holotype consists of a partial mandible. |
|||
|The smallest anthracothere from the Jebel Qatrani Formation and not as common than ''Bothriogenys''. |
|||
| |
|||
|- |
|||
|} |
|||
'''Chiroptera''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Dhofarella]]''<ref name=bats>{{Cite journal|last1=Simmons|first1=N.B.|last2=Seiffert|first2=E.R.|last3= Gunnell|first3=G.F.|year=2016|title=A new family of large omnivorous bats (Mammalia, Chiroptera) from the Late Eocene of the Fayum Depression, Egypt, with comments on use of the name “Eochiroptera”|url=https://digitallibrary.amnh.org/bitstream/handle/2246/6651/N3857%20High-res.pdf?sequence=2|journal=American Museum Novitates|issue=3857|pages=1-43|issn=0003-0082}}</ref><ref name=bat2/> |
|||
|''Dhofarella sigei'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A left dentary. |
|||
|A type of [[Emballonuridae|sheath-tailed bat]]. |
|||
| |
|||
|- |
|||
|''[[Khonsunycteris]]''<ref name=bats/> |
|||
|''Khonsunycteris aegypticus'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A left dentary. |
|||
|A type of [[Vespertilionidae|vesper bat]]. |
|||
| |
|||
|- |
|||
|[[Phasmatonycteris]]<ref name=sucker>{{Cite journal|last1=Gunnell|first1=G.F.|last2=Simmons|first2=N.B.|last3=Seiffert|first3=E.R.|year=2014|title=New Myzopodidae (Chiroptera) from the Late Paleogene of Egypt: Emended Family Diagnosis and Biogeographic Origins of Noctilionoidea|url=https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0086712|journal=PLoS ONE |volume=9|issue=2|doi=10.1371/journal.pone.0086712}}</ref><ref name=bats/> |
|||
|''Phasmatonycteris phiomensis'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|Dentary remains. |
|||
|A type of [[Myzopodidae|sucker-footed bat]]. |
|||
| |
|||
|- |
|||
|''[[Philisis]]''<ref name=bats/><ref name=bat2>{{Cite journal|last1=Gunnell|first1= G. F.|last2=Simons|first2=E. L.|last3=Seiffert|first3=E. R.|year=2008|title= New bats (Mammalia: Chiroptera) from the late Eocene and early Oligocene, Fayum Depression, Egypt|journal=Journal of Vertebrate Paleontology|volume=28|issue=1|pages=1–11|doi=10.1671/0272-4634(2008)28[1:nbmcft]2.0.co;2}}</ref> |
|||
|''Philisis sphingis'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|Material of the maxilla and dentary. |
|||
|A [[Philisidae|philisid]] bat related to the older ''[[Witwatia]]'' from Birket Qarun. |
|||
| |
|||
|- |
|||
|''[[Saharaderma]]''<ref name=bats/> |
|||
|''Saharaderma pseudovampyrus'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A right dentary. |
|||
|A type of [[Megadermatidae|false vampire bat]]. |
|||
| |
|||
|- |
|||
|[[Vampyravus]]<ref name=bats/><ref name=bat2/> |
|||
|''Vampyravus orientalis'' |
|||
|Upper Sequence |
|||
| |
|||
|A large humerus. |
|||
|A bat of uncertain affinities. It was the first fossil bat discovered in Africa and the largest bat of the Fayum succession, weighing up to {{convert|120|g|abbr=on}}. This puts it within the size range of the [[Egyptian fruit bat]]. |
|||
| |
|||
|- |
|||
|} |
|||
'''Embrithopoda''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Arsinoitherium]]''<ref name=C90>{{Cite journal|last1=Court|first1=N.|year=1990|title=Periotic anatomy of Arsinoitherium (Mammalia, Embrithopoda) and its phylogenetic implications|journal=Journal of Vertebrate Paleontology|volume=10|issue=2|pages=170–182|doi=10.1080/02724634.1990.10011806}}</ref><ref name=V13>{{Cite journal|last1=Vialle|first1=N.|last2=Merzeraud|first2=G.|last3=Delmer| first3=C.|last4=Feist|first4=M.|last5=Jiquel|first5=S.|last6=Marivaux|first6=L.|last7=Ramdarshan|first7=A.|last8=Vianey-Liaud|first8=M.|last9=Essid|first9=E.M.|last10=Marzougui|first10=W.|last11=Ammar| first11=H.K.|last12=Tabuce|first12=R.|year=2013|title=Discovery of an Embrithopod Mammal (Arsinoitherium?) in the Late Eocene of Tunisia.|url=https://d1wqtxts1xzle7.cloudfront.net/47397533/Discovery_of_an_embrithopod_mammal_Arsi20160721-24129-iajq0t-with-cover-page-v2.pdf?Expires=1667583660&Signature=K-gl-bkyQpAXgNI5G9vHtR3SMFjkfLJ4QYRWcnxVrD~BQvfoSUC7j44m6Bqm2fl8Gb7cK6DGiDwUy8-qvG6Q9CG8ilML7TDS3KMGT7zkMFelcGPe7SEmdipZ2LpfKOx6Rtj1KiCahGyDk1oAMfahxn21eNhcmmB6oZ9clIwjE7TYq-c-WWfOtesIpKrVFWRQnSyAwQvp3U9pt7XWnUuKrArzm5qi104eWVOEVMo9O1P1jgckWMG1O2duEcEhJVwc5NGoPNQ4Zcyw-VnM7d2QjxxqQjAfO55JlBMB-vVR2JIOuzr-wRaIwBOlWxlFIq83UX0OPPWam4p~qjavp91FrA__&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA|journal=Journal of African Earth Sciences|volume=87|pages=86–92|doi=10.1016/j.jafrearsci.2013.07.010}}</ref> |
|||
|''Arsinoitherium zitteli'' |
|||
|Lower and Upper Sequence |
|||
|Just above Quarry L-41 to Quarry M, spanning most of the sequence. |
|||
| |
|||
|A large [[Arsinoitheriidae|arsinoitheriid]] [[Embrithopoda|embrithopod]] well known for its two horns. A large amount of material is known and in the past a second species from Jebel Qatrani, ''A. andrewsi'', has been proposed. Some researchers however consider it to be a synonym of ''A. zitteli'' with the differences possibly being caused by sexual dimorphism. |
|||
|[[File:Arsinoitherium zitteli skull.JPG|frameless|center|''Arsinoitherium zitteli'' skull]] |
|||
|- |
|||
|} |
|||
'''Hyaenodonta''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|rowspan=2| |
|||
''[[Akhnatenavus]]''<ref name=BHS16/><ref name=O09/><ref name=Holroyd99/> |
|||
|''Akhnatenavus leptognathus'' |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|A slender mandible. |
|||
|A member of the [[Hyainailourinae]], it was originally described as ''Pterodon leptognathus''. It was smaller than the two other ''"Pterodon"'' species from Jebel Qatrani and ''Metapterodon''. |
|||
|[[File:Pterodon leptognathus.jpg|frameless|center]] |
|||
|- |
|||
|''Akhnatenavus nefertiticyon'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A complete cranium as well as a palate and dentary. |
|||
|A second species of ''Akhnatenavus'' smaller than ''A. leptognathus''. The skull also appears to have been much shorter than in the type species. Calculations resulted in an average bodymass of {{convert|19.2|kg|abbr=on}}, approximately as large as a [[Eurasian lynx]] or [[wolverine]]. |
|||
|[[File:Akhnatenavus skull Borths et al. 2016.png|frameless|center]] |
|||
|- |
|||
|rowspan=2| |
|||
''[[Apterodon]]''<ref name=O09>{{Cite journal|last1=Osborn|first1=H.F.|year=1909|title=New carnivorous mammals from the Fayûm Oligocene, Egypt. |url=https://digitallibrary.amnh.org/bitstream/handle/2246/1452//v2/dspace/ingest/pdfSource/bul/B026a28.pdf?sequence=1&isAllowed=y|journal=Bulletin of the American Museum of Natural History|volume=26| pages=415-424}}</ref><ref name=H96>{{Cite journal|last1=Holroyd|first1=P.A.|last2=Simons|first2=E.L.|last3=Bown|first3=T.M.|last4=Polly|first4=P.D.|last5=Kraus|first5=M.J.|year=1996|title=New records of terrestrial mammals from the upper Eocene Qasr el Sagha Formation, Fayum Depression, Egypt|url= https://pollylab.indiana.edu/doc/papers/Holroyd%20et%20al.,%20Qasr%20El%20Sagha.pdf |journal= Palaeovertebrata|volume=25|issue=2-4|pages=175-192}}</ref><ref name=K23/><ref name=SP76>{{Cite journal|last1=Simons|first1=E.L.|last2=Gingerich|first2=P.D.|year=1976|title=A new species of Apterodon (Mammalia, Creodonta) from the upper Eocene Qasr el-Sagha Formation of Egypt.|url=https://elischolar.library.yale.edu/cgi/viewcontent.cgi?article=1167&context=peabody_museum_natural_history_postilla|journal=Postilla of the Peabody Museum of Natural History}}</ref><ref name=LBB05>{{Cite journal|last1=Lange-Badré|first1=B.|last2=Böhme|first2=M.|year=2005|title=Apterodon intermedius, sp. nov., a new European creodont mammal from MP22 of Espenhain (Germany).|url=https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=7f96fdedeb3bcb3afa5fdf60efe60a915a06fbf5|journal=Annales de Paléontologie|volume=91|issue=4|doi=10.1016/j.annpal.2005.08.001}}</ref> |
|||
|''Apterodon altidens'' |
|||
|Lower Sequence |
|||
| |
|||
|Described based on a maxilla. |
|||
|Although named in 1910, ''A. altidens'' was not properly described until 1911. A mandible was also referred to this species, but this decision has been considered dubious by Lange-Badré and Böhme. |
|||
| |
|||
|- |
|||
|''Apterodon macrognathus'' |
|||
|Lower Sequence |
|||
|Quarry A & B |
|||
|Skulls and mandibles |
|||
|A hyaenodont originally described as a species of ''[[Pterodon]]''. |
|||
|[[File:Apterodon macrognathus skull2.svg|frameless|center]] |
|||
|- |
|||
|''[[Brychotherium]]''<ref name=BHS16/><ref name=BS17/> |
|||
|''Brychotherium ephalmos'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Multiple lower jaws and parts of the rostrum. |
|||
|A teratodontine hyaenodont approximately as heavy as a modern red fox or American badger. Although the name was coined in a dissertation in 1994, it was not formally described until 2016. |
|||
|[[File:Brychotherium dentary Holotype Borths et al 2016.png|frameless|center]] |
|||
|- |
|||
|''[[Falcatodon]]''<ref name=BS17b/><ref name=MP17/><ref name=Holroyd99>{{Cite journal|last1=Holroyd|first1=P.A.|year=1999|title=New Pterodontinae (Creodonta: Hyaenodontidae) from the late Eocene-early Oligocene Jebel Qatrani Formation, Fayum province, Egypt.|url=https://docubase.berkeley.edu/cgi-bin/pl_dochome?query_src=pl_search&collection=PaleoBios+Archive+Public&id=141|journal=PaleoBios|volume=19|issue=2|pages=1-18}}</ref> |
|||
|''Falcatodon schlosseri'' |
|||
|Upper Sequence |
|||
|Quarry V |
|||
|Described based on a left dentary. |
|||
|Erected by Holroyd as a species of ''Metapterodon'', it was even then noted to be possibly distinct. Morales and Pickford later raised it to a distinct genus. There may be additional material from Quarry B in the lower sequence, however the assignment of said fossils is only tentative. |
|||
|[[File:Falcatodon 2.jpg|frameless|center]] |
|||
|- |
|||
|rowspan=2| |
|||
''[[Masrasector]]''<ref name=K23/><ref name=BHS16/><ref name=BS17>{{Cite journal|last1=Borths|first1=M.R.|last2=Seiffert|first2=E.R.|year=2017|title=Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts|url=https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0173527|journal=PLoS ONE|volume=12|issue=4|doi=10.1371/journal.pone.0173527}}</ref> |
|||
|''Masrasector aegypticum'' |
|||
|Upper Sequence |
|||
|Quarry G |
|||
|Dental remains |
|||
|A teratodontine hyaenodont smaller than ''Brychotherium''. |
|||
| |
|||
|- |
|||
|''Masrasector nananubis'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Skulls, mandibles and multiple [[humeri]]. |
|||
|A smaller species of ''Masrasector'' from older deposits. It's estimated boy size was within the range of modern [[striped skunk]]s and [[small-spotted genet]]s. The limb bones indicate it was a fast runner that could have hunted the local rodents. |
|||
|[[File:Masrasector cranium2 Borths et al 2017.tif|frameless|center]] |
|||
|- |
|||
|''[[Metapterodon]]''<ref name=O09/><ref name=Holroyd99/><ref name=H96/> |
|||
|''Metapterodon brachycephalus'' |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|A left dentary. |
|||
|Named as a species of ''Hyaenodon'', it was eventually placed in ''Metapterodon'' by Holroyd. While Morales and Pickford restrict the genus to the Miocene and place Holroyd's other species in separate genera, they do not mention ''M. brachycephalus''. |
|||
|[[File:Hyaenodon brachycephalus.jpg|frameless|center]] |
|||
|- |
|||
|''[[Metasinopa]]''<ref name=O09/><ref name=BHS16/><ref name=BS17/> |
|||
|''Metasinopa fraasi'' |
|||
|Upper Sequence |
|||
| |
|||
|A nearly complete lower jaw. |
|||
|A possible relative to teratodontines, although its exact relationship with other hyaenodontids needs more testing. It was larger than ''Masrasector''. |
|||
|[[File:Metasinopa fraasii.jpg|frameless|center]] |
|||
|- |
|||
|rowspan=4| |
|||
''"[[Pterodon (mammal)|Pterodon]]"''<ref name=BHS16/><ref name=K23/><ref name=O09/><ref name=Holroyd99/><ref name=BS17b/> |
|||
|''[["Pterodon" africanus]]'' |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|Mandibles, a rostrum and an assigned neck vertebra. |
|||
|The first hyaenodont described from Fayum, later analysis showed that it was much more derived than ''Pterodon dasyuroides'', the type species. Consequently, a different genus name is required. |
|||
|[[File:Pterodon africanus.jpg|frameless|center]] |
|||
|- |
|||
|''[["Pterodon" phiomensis]]'' |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|A mandible smaller than that of ''"P." africanus''. |
|||
|Like ''"P." africanus'', ''"P." phiomensis'' is much more derived than the type species of ''Pterodon'' and thus requires a different genus name. Furthermore, analysis showed that it was not closely related to ''"P." africanum'' either. It is most closely allied with the clade formed by ''Akhnatenavus'', ''[[Isohyaenodon]]'' and ''[[Hyainailouros]]''. |
|||
|[[File:Pterodon phiomensis jaw.svg|frameless|center]] |
|||
|- |
|||
|''"Pterodon" sp.'' |
|||
|Upper Sequence |
|||
|Quarry E |
|||
|A broken canine. |
|||
|An isolated tooth similar in size to ''"P." phiomensis''. |
|||
| |
|||
|- |
|||
|''[["Pterodon" syrtos]]'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A right maxilla fragment. |
|||
| |
|||
| |
|||
|- |
|||
|''[[Quasiapterodon]]''<ref name=LBB05/><ref name=K23/><ref name=BS17b>{{Cite journal|last1=Borths|first1=M.R.|last2=Stevens|first2=N.J.|year=2017|title=The first hyaenodont from the late Oligocene Nsungwe Formation of Tanzania: Paleoecological insights into the Paleogene-Neogene carnivore transition|url=https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0185301|journal=PLoS ONE|volume=12|issue=10|doi=10.1371/journal.pone.0185301}}</ref><ref name=S14>{{Cite journal|last1=Solé|first1=F.|last2=Lhuillier|first2=J.|last3=Adaci|first3=M.|last4=Bensalah|first4=M.|last5=Mahboubi|first5=M.|last6=Tabuce|first6=R.|year=2014|title=The hyaenodontidans from the Gour Lazib area (? early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae|url=https://d1wqtxts1xzle7.cloudfront.net/45533449/The_hyaenodontids_from_the_Gour_Lazib_ar20160511-22943-txg8mz-libre.pdf?1462957861=&response-content-disposition=inline%3B+filename%3DThe_hyaenodontidans_from_the_Gour_Lazib.pdf&Expires=1683628185&Signature=TcY~JcXycn6X1wlvmBW~mWV087eGjC30JWg-NbZ9Ff2Sr4SmorVCvGPkJRlPrwwCUMmqXWuVRhcf1v-ycMUrT2NnmbQg85QLqg0ILCula4x~oxE9oAfGzJBPkvL2~g0iA9F6qdu4V98WxL3ANk9RO4PPWf7mCfffdEiocYQtkSpL0nm1d~g3~VznukGzBPOr3c5yGL91i68rwcBAesHam8bTcz4cF16GuhXgdEu9TSlZ25b11RwYx2Ju3zqTqDkg2DweK9fvPd508k3e9sag524MuIDJeMcMYux4INrv8fbxU8YieGLZtuT2mFKpog77kyGXB9x3lz7LgNjxzUX23g__&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA|journal=Journal of Systematic Palaeontology |volume=12|issue=3|pages=303-322|doi=10.1080/14772019.2013.795196}}</ref> |
|||
|''Quasiapterodon minutus'' |
|||
|Upper Sequence |
|||
| |
|||
| |
|||
|Originally named as a species of ''Apterodon'' alongside ''A. altidens'', later research showed it clearly differed from this genus. It may instead be related to ''"Sinope" ethiopica''. |
|||
| |
|||
|- |
|||
|''[[Sectisodon]]''<ref name=BS17b/><ref name=MP17>{{Cite journal|last1=Morales|first1=J.|last2=Pickford|first2=M.|year=2017|title=New hyaenodonts (Ferae, Mammalia) from the early Miocene of Napak (Uganda), Koru (Kenya) and Grillental (Namibia)|journal=Fossil Imprint|volume= 73|issue=3–4|pages=332–359|doi=10.2478/if-2017-0019}}</ref><ref name=Holroyd99/> |
|||
|''Sectisodon markgrafi'' |
|||
| |
|||
| |
|||
|A left maxilla. |
|||
|''S. markgrafi'' was initially named by Holroyd as a species of ''Metapterodon'' in 1999, but placed in the new genus ''Sectisodon'' by Morales and Pickford in 2017. |
|||
| |
|||
|- |
|||
|''"[[Sinopa]]"''<ref name=Sino>{{Cite journal|last1=Andrews|first1=C.W.|year=1906|title=A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt|url= https://www.biodiversitylibrary.org/page/37623110#page/278/mode/1up | journal=Order of the Trustees of the British Museum}}</ref><ref name=BHS16>{{Cite journal|last1=Borths|first1=M.R.|last2=Holroyd|first2=P.A.|last3=Seiffert| first3=E.R.|year=2016|title=Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia)|url=https://peerj.com/articles/2639/|journal=PeerJ|doi=10.7717/peerj.2639}}</ref><ref name=LBB05/><ref name=MM>{{Cite journal|last1=Morlo|first1=M.|last2=Bastl|first2=K.|last3=Wenhao|first3=W.|last4=Schaal |first4=S.F.K. | year=2014|title=The first species of Sinopa (Hyaenodontida, Mammalia) from outside of North America: implications for the history of the genus in the Eocene of Asia and North America.|url=https://onlinelibrary.wiley.com/doi/pdf/10.1111/pala.12052|journal=Palaeontology|volume=57|issue=1|pages=111-125|doi=10.1111/pala.12052}}</ref> |
|||
|''[["Sinopa" ethiopica]]'' |
|||
| |
|||
| |
|||
|A mandibular ramus. |
|||
|A hyaenodont originally described as a species of ''Sinopa'', although later research clearly shows that it does not belong to the American-Asian genus. It's exact relation to other hyaenodonts remains uncertain until further study, however it has been suggested to be a relative of ''Quasiapterodon''. |
|||
| |
|||
|- |
|||
|} |
|||
Holroyd also identified several indetermined pterodontine hyaenodonts from various sequences of the Jebel Qatrani Formation, but doesn't identify them beyond subfamily level.<ref name=Holroyd99/> |
|||
'''Hyracoidea''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Antilohyrax]]''<ref name=RS2000>{{Cite journal|last1=Rasmussen|first1=D.T.|last2=Simons|first2=E.L.|year=2000|title=Ecomorphological diversity among Paleogene hyracoids (Mammalia): a new cursorial browser from the Fayum, Egypt.|url=https://www.researchgate.net/profile/David-Rasmussen-4/publication/232686389_Ecomorphological_diversity_among_Paleogene_hyracoids_Mammalia_A_new_cursorial_browser_from_the_Fayum_Egypt/links/0912f5109593855d63000000/Ecomorphological-diversity-among-Paleogene-hyracoids-Mammalia-A-new-cursorial-browser-from-the-Fayum-Egypt.pdf|journal=Journal of Vertebrate Paleontology|volume=20|issue=1|pages=167-176|doi=10.1671/0272-4634(2000)020[0167:EDAPHM]2.0.CO;2}}</ref><ref name=DB02>{{Cite journal|last1=De Blieux| first1=D.D.|last2=Simons|first2=E.L.|year=2002|title=Cranial and dental anatomy of Antilohyrax pectidens: a late Eocene hyracoid (Mammalia) from the Fayum, Egypt.|journal=Journal of Vertebrate Paleontology|volume=22|issue=1 |pages=122-136|doi=10.1671/0272-4634(2002)022[0122:CADAOA]2.0.CO;2}}</ref> |
|||
|''Antilohyrax pectidens'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Mandibles, a crushed cranium as well as various incomplete skull fossils and postcranial elements. |
|||
|A gazelle-like titanohyracine described in 2000. The teeth and limb bones indicate that it was a cursorial browser, similar to modern antelopes and gazelles. |
|||
| |
|||
|- |
|||
|rowspan=3| |
|||
''[[Bunohyrax]]'' |
|||
|''Bunohyrax fajumensis''<ref name=DeBlieux/><ref name=M73/><ref name=Tabu/> |
|||
|Lower and Upper Sequence |
|||
| |
|||
|Described based on a mandible. |
|||
|''Bunohyrax'' species were medium-sized hyracoids somewhat resembling pigs in their dentition. They may have been related to ''Pachyhyrax''. |
|||
| |
|||
|- |
|||
|''Bunohyrax major''<ref name=DeBlieux/><ref name=Tabu/> |
|||
|Upper Sequence |
|||
| |
|||
|Described based on three premolars. |
|||
|Originally named ''Geniohyus major'', ''B. major'' is larger than ''B. fajumensis''. Additionally, its teeth are more bunodont. |
|||
| |
|||
|- |
|||
|''Bunohyrax sp.''<ref name=RS2000/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|rowspan=4| |
|||
''[[Geniohyus]]''<ref name=K23/> |
|||
|''Geniohyus diphycus''<ref name=RS88/> |
|||
|Lower Sequence |
|||
| |
|||
| |
|||
|A medium-sized species. ''Geniohyus'' is among the rarer hyracoids of Jebel Qatrani. |
|||
| |
|||
|- |
|||
|''Geniohyus magnus''<ref name=Tabu/><ref name=RS88/> |
|||
|Lower and Upper Sequence |
|||
|Quarry V |
|||
|Various skull remains including maxillary fragments and dentary remains. |
|||
|It is the smallest species of ''Geniohyus'', with the name originating when it was still considered apart of ''Saghatherium''. It's dentition forms an intermediate between other ''Geniohyus'' and ''Bunohyrax''. |
|||
| |
|||
|- |
|||
|''Geniohyus mirus''<ref name=DeBlieux/><ref name=Tabu/><ref name=RS88/> |
|||
|Lower Sequence |
|||
| |
|||
|Described based on a mandible. |
|||
|The largest of the three named Fayum ''Geniohyus'' species and the type species. |
|||
| |
|||
|- |
|||
|''Geniohyus sp.''<ref name=RS2000/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|''Geniohyus'' takes the species name from their morphological similarities to pigs, with early research even classifying them as [[Suidae|suids]]. |
|||
| |
|||
|- |
|||
|rowspan=2| |
|||
''[[Megalohyrax]]''<ref name=M22>{{Cite journal|last1=Matsumoto|first1=H.|year=1922|title=Megalohyrax, Andrews and Titanohyrax, gen. nov. A revision of the genera of hyracoids from the Fayum, Egypt|journal=Proceedings of the Zoological Society|issue=1921|pages=839-850}}</ref> |
|||
|''Megalohyrax eocaenus''<ref name=RS2000/><ref name=DeBlieux/><ref name=M73/><ref name=Tabu/> |
|||
|Upper Sequence |
|||
| |
|||
|Described based on premaxillary and maxillary material. Also includes mandibular remains. |
|||
|''Megalohyrax'' show strong sexual dimorphism in regards to their body size. It was a common genus in formation. |
|||
| |
|||
|- |
|||
|''Megalohyrax sp.''<ref name=RS2000/><ref name=DeBlieux/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|An unnamed species of ''Megalohyrax'' preserving an interior mandibular fenestra unlike the younger ''M. eocaenus''. |
|||
| |
|||
|- |
|||
|''[[Pachyhyrax]]''<ref name=RS2000/><ref name=DeBlieux/><ref name=M73/><ref name=RS88/> |
|||
|''Pachyhyrax crassidentatus'' |
|||
|Upper Sequence |
|||
|Quarry I, L-46 & M |
|||
|Skulls, mandibles and teeth including the complete but crushed skull of a young individual. |
|||
|A hyracoid resembling the contemporary anthracothere ''Bothriogenys''. It is possible that it was a semi-aquatic herbivore. |
|||
| |
|||
|- |
|||
|rowspan=3| |
|||
''[[Saghatherium]]''<ref name=M73/> |
|||
|''Saghatherium antiquum''<ref name=DeBlieux/><ref name=RTE91/><ref name=Tabu/><ref name=RS88/> |
|||
|Lower Sequence |
|||
| |
|||
|Described based on a maxilla |
|||
|''Saghatherium'' had heavy jaws and possibly fed on nuts and seeds. Furthermore, members of this genus display pronounced [[sexual dimorphism]] and may have had complex mating rituals. The species ''S. sobrina'' was shown to simply represent female individuals of this speices. |
|||
|rowspan=3| |
|||
[[File:Saghatherium antiquum.JPG|frameless|center]] |
|||
|- |
|||
|''Sagahtherium humarum''<ref name=RS88/> |
|||
|Upper Sequence |
|||
|Quarry V |
|||
|Dentaries and teeth. |
|||
|A small ''Saghatherium'' species that differs due to the presence of an internal chamber in the mandible. |
|||
|- |
|||
|''Saghatherium bowni''<ref name=RS2000/><ref name=DeBlieux/><ref name=RTE91/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|A common species of ''Saghatherium'' named in 1991. |
|||
|- |
|||
|''[[Selenohyrax]]<ref name=RS2000/><ref name=DeBlieux/><ref name=RS88>{{Cite journal|last1=Rasmussen|first1=D. T.|last2=Simons|first2=E. L.|year=1988|title=New Oligocene hyracoids from Egypt|journal=Journal of Vertebrate Paleontology|volume=8|issue=1|pages=67–83|doi=10.1080/02724634.1988.10011684}}</ref> |
|||
|''Selenohyrax chatrathi'' |
|||
|Upper Sequence |
|||
|Quarry V |
|||
|Several mandibles. |
|||
|A hyracoid named for its selenodont teeth (molars with crescent-shaped ridges), which are much more developed than in ''Titanohyrax''. These teeth indicate a diet that required it to slice vegetation rather than grind it. It is a small hyrax, only slightly larger than ''Saghatherium'' which it might have descendet from. |
|||
| |
|||
|- |
|||
|rowspan=4| |
|||
''[[Thyrohyrax]]'' |
|||
|''Thyrohyrax domorictus''<ref name=DeBlieux/><ref name=M73>{{Cite journal|last1=Meyer|first1=G.E.|year=1973|title=A new Oligocene hyrax from the Jebel el Qatrani formation, Fayum, Egypt| url=https://ia800207.us.archive.org/12/items/biostor-118529/biostor-118529.pdf |journal=Peabody Museum of Natural History, Yale University}}</ref> |
|||
|Upper Sequence |
|||
|Quarry I, G, L-46, M, P, R, V & X |
|||
|Primarily mandibular remains and teeth, but also some limb elements and vertebrae. |
|||
|''Thyrohyrax'' is among the smaller hyracoids of Fayum and resembles today's arboreal ''[[Dendrohyrax]]''. Species of ''Thyrohyrax'' display little size-related sexual dimorphism, however males do possess enlarged, tusk-like second [[incisors]] and chambered mandibles. |
|||
| |
|||
|- |
|||
|''Thyrohyrax litholagus''<ref name=RS2000/><ref name=RTE91>{{Cite journal|last1=Rasmussen|first1=D.T.|last2=Simons|first2=E.L.|year=1991|title=The Oldest Egyptian Hyracoids (Mammalia: Pliohyracidae): New Species of Saghatherium and Thyrohyrax from the Fayum|journal=Neues Jahrbuch für Geologie und Paläontologie|volume=182|issue=2|pages=187 - 209|doi=10.1127/njgpa/182/1991/187}}</ref> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|A relative of the younger ''Thyrohyrax pygmaeus'', it was somewhat larger than ''T. meyeri''. |
|||
| |
|||
|- |
|||
|''Thyrohyrax meyeri''<ref name=RS2000/><ref name=DeBlieux>{{Cite journal|last1=De Blieux|first1=D.D.|last2=Baumrind|first2=M.R.|last3=Simons|first3=E.L.|last4=Chatrath|first4=P.S.|last5=Meyer|first5=G.E.|last6=Attia|first6=Y.S.| year=2006|title=Sexual dimorphism of the internal mandibular chamber in Fayum Pliohyracidae (Mammalia).|url=https://www.researchgate.net/profile/Donald-Deblieux/publication/228910875_Sexual_dimorphism_of_the_internal_mandibular_chamber_in_Fayum_Pliohyracidae_Mammalia/links/5668533c08ae8d6928fa7e47/Sexual-dimorphism-of-the-internal-mandibular-chamber-in-Fayum-Pliohyracidae-Mammalia.pdf|journal=Journal of Vertebrate Paleontology|volume=26|issue=1|pages=160-169}}</ref><ref name=RTE91/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|''T. meyeri'' was very similar to the younger ''T. domorictus'' and may have been a close relative. |
|||
| |
|||
|- |
|||
|''Thyrohyrax pygmaeus''<ref name=DeBlieux/><ref name=RS88/> |
|||
|Lower Sequence |
|||
|Quarry A-1 |
|||
|The front of a skull with an associated mandible as well as several teeth from another specimen. |
|||
|A close relative of direct descendent of ''T. litholagus''. It was originally assigned to ''Saghatherium magnum'', then described as a species of ''Megalohyrax'' before being moved into ''Pachyhyrax'' and eventually ''Thyrohyrax''. |
|||
| |
|||
|- |
|||
|rowspan=4| |
|||
''[[Titanohyrax]]''<ref name=M22/> |
|||
|''Titanohyrax andrewsi''<ref name=RS2000/><ref name=Tabu>{{Cite journal|last1=Tabuce|first1=R.|year=2016|title=A mandible of the hyracoid mammal Titanohyrax andrewsi in the collections of the Muséum National d'Histoire Naturelle, Paris (France) with a reassessment of the species.|url=https://hal.science/hal-03100188/document|journal=Palaeovertebrata|volume=40|issue=1|doi=10.18563/pv.40.1.e4}}</ref><ref name=RS88/> |
|||
|Lower Sequence |
|||
| |
|||
|Five specimen including two mandibles and a single maxilla. |
|||
|''Titanohyrax'' species are the largest hyracoids of the Fayum Depression and form the clade [[Titanohyracinae]] with ''Antilohyrax''. Like their smaller relative, they are thought to have been folivores. The lower incisors of ''T. andrewsi'' were shaped like spatulas, rather than tusks. |
|||
| |
|||
|- |
|||
|''Titanohyrax angustidens''<ref name=RS2000/><ref name=Tabu/><ref name=RS88/> |
|||
|Upper Sequence |
|||
|Quarry V, R & I |
|||
|Eight specimen including maxillary, premaxillary and mandibular remains. |
|||
|''T. angustidens'' has the most complex history among ''Titanohyrax species'', initially assigned to ''Megalohyrax'' and given the name ''M. palaeotheroides'', it was later recognized as ''Titanohyrax'' and variably considered distinct or synonymous with ''T. andrewsi''. The name ''T. palaeotheroides'' was not retained as it was a [[nomen nudum]]. |
|||
| |
|||
|- |
|||
|''Titanohyrax ultimus''<ref name=RS2000/><ref name=Tabu/> |
|||
|Upper Sequence |
|||
| |
|||
|Four heavily worn teeth. |
|||
|The largest species of ''Titanohyrax'', weight estimates for ''T. ultimus'' range from {{convert|600-1300|kg|abbr=on}}, making it possibly as large as the modern [[sumatran rhino]]. |
|||
| |
|||
|- |
|||
|''Titanohyrax sp.''<ref name=RS2000/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|An unnamed species from the L-41 Quarry. |
|||
| |
|||
|- |
|||
|} |
|||
'''Marsupalia''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Peratherium]]''<ref name=K23/><ref name=Pera>{{Cite journal|last1=Hooker|first1=J.J.|last2=Sánchez-Villagra|first2=M.R.|last3=Goin|first3=F.J.|last4=Simons|first4=E.L.|last5=Attia|first5=Y.|last6=Seiffert|first6=E.R.| year=2008|title=The origin of Afro‐Arabian ‘didelphimorph’marsupials|url=https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.1475-4983.2008.00779.x|journal=Palaeontology|volume=51|issue=3|pages=635-648|doi=10.1111/j.1475-4983.2008.00779.x}}</ref> |
|||
|''Peratherium africanum'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|Several specimen including a maxilla and three mandibles |
|||
|A primitive [[marsupial]] and the basalmost mammal found in the sediments of the formation. It was at one point known under the name ''Qatranitherium'', however this name has since then been deemed a junior synonym. |
|||
| |
|||
|- |
|||
|} |
|||
'''Rodentia''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|rowspan=3 | |
|||
''[[Acritophiomys]]''<ref name=AA21/><ref name=SSS12>{{Cite journal|last1=Sallam|first1=H.M.|last2=Seiffert|first2=E.R.|last3=Simons|first3= E.L. |year=2012|title=A basal phiomorph (Rodentia, Hystricognathi) from the late Eocene of the Fayum Depression, Egypt|url=https://sjpp.springeropen.com/articles/10.1007/s13358-012-0039-6|journal=Swiss Journal of Palaeontology|volume= 131|pages=283–301|doi=10.1007/s13358-012-0039-6}}</ref><ref name=BS17/> |
|||
|''Acritophiomys adaios'' |
|||
|Lower Sequence |
|||
|Quarry A, B & E |
|||
| |
|||
|This species also includes several specimen previously assigned to ''Phiomys andrewsi''. However, as it was not adequately described it is considered a [[nomen nudum]]. |
|||
| |
|||
|- |
|||
|''Acritophiomys bowni'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Complete upper and lower dentition as well as parts of the lower jaw and the skull. |
|||
|A basal member of [[Phiomorpha]] and the largest member of the family "[[Phiomyidae]]", although the group is now considered to be [[paraphyletic]]. It is the only species of ''Acritophiomys'' that had been validly named, as both ''A. adaios'' and ''A. woodi'' were only described in a [[thesis]]. |
|||
| |
|||
|- |
|||
|''Acritophiomys woodi'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|The first phiomydid described from the Quarry L-41. However, as it was not adequately described it is considered a nomen nudum. |
|||
| |
|||
|- |
|||
|''[[Birkamys]]''<ref name=AA21/><ref name=K23/><ref name=SaSe16>{{Cite journal|last1=Sallam|first1=H.M.|last2=Seiffert|first2=E.R.|year=2016|title=New phiomorph rodents from the latest Eocene of Egypt, and the impact of Bayesian “clock”-based phylogenetic methods on estimates of basal hystricognath relationships and biochronology|url=https://peerj.com/articles/1717/|journal=PeerJ|volume=4|doi=10.7717/peerj.1717}}</ref> |
|||
|''Birkamys korai'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Cranial and mandibular remains with teeth, including a partially preserved rostrum. |
|||
|A close relative of ''Mubhammys''. |
|||
| |
|||
|- |
|||
|rowspan=3| |
|||
''[[Gaudeamus (rodent)|Gaudeamus]]''<ref name=AA21/><ref name=K23/><ref name=SSS12/><ref name=SaSeSi11>{{Cite journal|last1=Sallam|first1=H.M.|last2=Seiffert|first2=E.R.|last3=Simons|first3=E.L.|year=2011|title=Craniodental Morphology and Systematics of a New Family of Hystricognathous Rodents (Gaudeamuridae) from the Late Eocene and Early Oligocene of Egypt|url=https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0016525|journal=PLoS ONE |volume=6|issue=2|doi=10.1371/journal.pone.0016525}}</ref><ref name=BS17/> |
|||
|''Gaudeamus aegyptius'' |
|||
|Upper Sequence |
|||
|Quarry E |
|||
|Mandibles and teeth |
|||
|The type species of the genus, described by Wood in the 1960s. It is intermediate in size among the Fayum rodents, larger than the small ''Phiocricetomys'' but smaller than ''Metaphiomys''. Its phylogeny is enigmatic and it has variably been placed as a sister to [[old world porcupines]] or within [[Caviomorpha]]. |
|||
| |
|||
|- |
|||
|''Gaudeamus aslius'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A distorted skull and various fragmentary remains. |
|||
|It shows several primitive features and may resemble the ancestral form of this genus. |
|||
| |
|||
|- |
|||
|''Gaudeamus hylaeus'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A flattened skull and various additional fragments. |
|||
|''G. hylaeus'' was initially named by Holroyd in a PhD Thesis, rendering it a nomen nudum until the full description authored by Sallam, Seiffert and Simons. |
|||
| |
|||
|- |
|||
|rowspan=2| |
|||
''[[Metaphiomys]]''<ref name=K23/><ref name=SSS12/> |
|||
|''Metaphiomys beadnelli'' |
|||
|Upper Sequence |
|||
| |
|||
|A left mandible and additional material. |
|||
|A phiomyid rodent. |
|||
| |
|||
|- |
|||
|''Metaphiomys schaubi'' |
|||
| |
|||
| |
|||
| |
|||
|A second species of ''Metaphiomys'' described by Wood in the 1960s. |
|||
| |
|||
|- |
|||
|''[[Monamys]]''<ref name=SSS12/><ref name=K23/><ref name=SaSe20>{{Cite journal|last1=Sallam|first1=H.M.|last2=Seiffert|first2=E.R.|year=2020|title=Revision of Oligocene ‘Paraphiomys’ and an origin for crown Thryonomyoidea (Rodentia: Hystricognathi: Phiomorpha) near the Oligocene–Miocene boundary in Africa|url=https://www.researchgate.net/profile/Erik-Seiffert/publication/338281768_Revision_of_Oligocene_'Paraphiomys'_and_an_origin_for_crown_Thryonomyoidea_Rodentia_Hystricognathi_Phiomorpha_near_the_Oligocene-Miocene_boundary_in_Africa/links/628a56608ecbaa07fccbb9b0/Revision-of-Oligocene-Paraphiomys-and-an-origin-for-crown-Thryonomyoidea-Rodentia-Hystricognathi-Phiomorpha-near-the-Oligocene-Miocene-boundary-in-Africa.pdf|journal=Zoological Journal of the Linnean Society |volume=190|issue=1|pages=352-371}}</ref> |
|||
|''Monamys simonsi'' |
|||
|Upper Sequence |
|||
|Quarry I & M |
|||
|Mandibular remains and teeth. |
|||
|Previously known as ''[[Paraphiomys]] simonsi'', later research has shown that this genus is distinct from the much later Miocene ''Paraphiomys''. |
|||
| |
|||
|- |
|||
|''[[Mubhammys]]''<ref name=K23/><ref name=SaSe16/> |
|||
|''Mubhammys vadumensis'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Cranial and mandibular elements with teeth. |
|||
|A close relative of ''Birkamys''. |
|||
| |
|||
|- |
|||
|''[[Phiocricetomys]]''<ref name=AA21>{{Cite journal|last1=Al-Ashqar|first1=SF|last2=Seiffert|first2=E.R.|last3=de Vries|first3=D.|last4=El-Sayed|first4=S.|last5=Antar|first5= M.S. |last6=Sallam|first6=H.M.|year=2021|title=New phiocricetomyine rodents (Hystricognathi) from the Jebel Qatrani Formation, Fayum Depression, Egypt|url=https://peerj.com/articles/12074/|journal=PeerJ|volume=9|doi=10.7717/peerj.12074}}</ref><ref name=SSS12/> |
|||
|''Phiocricetomys minutus'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|A mandible preserving dentition from the 4th deciduous premolar to 2nd molar. |
|||
|The youngest known member of [[Phiocricetomyinae]]. |
|||
| |
|||
|- |
|||
|rowspan=2| |
|||
''[[Phiomys]]''<ref name=K23/><ref name=SSS12/> |
|||
|''Phiomys andrewsi'' |
|||
|Lower Sequence |
|||
|Quarry A & B |
|||
|Described based on partial mandibles. |
|||
|A phiomyid rodent. |
|||
| |
|||
|- |
|||
|''Phiomys paraphiomyoides'' |
|||
| |
|||
| |
|||
| |
|||
|A phiomyid described by Wood in the 1960s. |
|||
| |
|||
|- |
|||
|''[[Qatranimys]]''<ref name=AA21/> |
|||
|''Qatranimys safroutus'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Skull remains and teeth. |
|||
|A diminutive phiocricetomyine rodent. |
|||
| |
|||
|- |
|||
|''[[Talahphiomys]]''<ref name=AA21/><ref name=SSS12/><ref name=JJJ10>{{Cite journal|last1=Jaeger|first1=J.J.|last2=Marivaux|first2=L.|last3=Salem|first3=M.|last4=Bilal|first4=A.A.|last5=Benammi |first5=M.| last6=Chaimanee |first6=Y.|last7=Duringer|first7=P.|last8=Marandat|first8=B.|last9=Métais|first9=E.|last10=Schuster|first10=M.|last11=Valentin|first11=X.|last12=Brunet|first12=M.|year=2010|title=New rodent assemblages from the Eocene Dur At-Talah escarpment (Sahara of central Libya): systematic, biochronological, and palaeobiogeographical implications.|url=https://d1wqtxts1xzle7.cloudfront.net/91533146/j.1096-3642.2009.00600.x-libre.pdf?1664128801=&response-content-disposition=inline%3B+filename%3DNew_rodent_assemblages_from_the_Eocene_D.pdf&Expires=1683538625&Signature=fqyOaehCWi5hpP7JLWrUj1fK70wE5r5ig4mXUjUWGlazx29tRM8qnRYzJDiFDNdfRHUV0aJ5d4e0teJGVp1dhuHpiK9Dm3z8uguTHYIU6cwRWSVdyspbwzftj2xgNAkl7XtVB0lksSBOPuTiCy2p5VW3Zs-EZUuQLYx48dNo~~54QinPWtyi49DjLQcckS0X9BiKyTb1jfGpXQztBbfDR-fmEFB0InA5izThi6o1XBK1DRYD0bmf69kSmnNaD6vbj2gzFWwywTW54UvRaq5-bMh7AGTy4binRFygEM75I1-o00oELciE9Akm6SssLRe79iNX6ix5g5HB0ITqr8MnGA__&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA|journal=Zoological Journal of the Linnean Society|volume=160|issue=1|pages=195-213}}</ref> |
|||
|''Talahphiomys lavocati'' |
|||
|Lower Sequence |
|||
|Quarry E |
|||
|Deciduous premolars and molars. |
|||
|Initially thought to be a species of ''[[Phiomys]]'', it was named ''Elwynomys'' in a doctoral dissertation from 1994. Due to the rules of the [[ICZN]], the name ''Talahphiomys'' later took precedence. |
|||
| |
|||
|- |
|||
|} |
|||
'''Pholidota''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|[[Pholidota]]<ref name=K23/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M & L-12 |
|||
|Two finger bones. |
|||
|The fragmentary remains of what may have been a pangolin. The presence of termite nests bearing excavation marks may support this, however the identity of these finger bones is not universally supported and has been questioned. |
|||
| |
|||
|- |
|||
|} |
|||
'''Ptolemaiida''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|rowspan=2 | |
|||
[[Cleopatrodon]]<ref name=B87>{{Cite journal|last1=Bown|first1=T.M.|last2=Simons|first2=E.L.|year=1987|title=New Oligocene Ptolemaiidae (Mammalia: ?Pantolesta) from the Jebel Qatrani Formation, Fayum Depression, Egypt|journal=Journal of Vertebrate Paleontology|volume=7|issue=3|pages=311-324|doi=10.1080/02724634.1987.10011662}}</ref><ref name=S95>{{Cite journal|last1=Simons|first1=E.L.|last2=Bown| first2=T.M.|year=1995|title= Ptolemaiida, a new order of Mammalia--with description of the cranium of Ptolemaia grangeri|journal=Proc Natl Acad Sci U S A.|volume=92|issue=8|pages=3269-73|doi= 10.1073/pnas.92.8.3269|pmid=11607526|pmc=42147}}</ref> |
|||
|''Cleopatrodon ayeshae'' |
|||
|Upper Sequence |
|||
|Quarry V |
|||
|Mandibular remains and teeth. |
|||
|A [[Ptolemaiidae|ptolemaiid]] afrothere. The type species of ''Cleopatrodon''. |
|||
| |
|||
|- |
|||
|''Cleopatrodon robusta'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|Mandibular remains and teeth. |
|||
|''C. robusta'' had more robust jaws than ''C. ayeshae''. |
|||
| |
|||
|- |
|||
|rowspan=2 | |
|||
[[Ptolemaia]]<ref name=S95/> |
|||
|''Ptolemaia grangeri''<ref name=B87/> |
|||
|Upper Sequence |
|||
|Quarry V |
|||
|Multiple teeth and a mandibular ramus as well as an almost complete but distorted skull. |
|||
|A carnivorous afrotherian mammal of the family Ptolemaiidae. It is larger than the older ''P. lyonsi''. |
|||
| |
|||
|- |
|||
|''Ptolemaia lyonsi''<ref name=Osborn1908>{{cite journal|last=Osborn|first= H. F.|year=1908|title=New fossil mammals from the Fayum Oligocene, Egypt|journal= Bulletin of the American Museum of Natural History|volume=24|pages=265–272|url=https://www.biodiversitylibrary.org/page/27046076#page/317/mode/1up }}</ref> |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|Lower jaw material. |
|||
|A carnivorous afrothere first described by Osborn in 1908. |
|||
|[[File:Ptolemaia lyonsi jaw (cropped).jpg|frameless|center|Ptolemaia lyonsi jaw]] |
|||
|- |
|||
|''[[Qarunavus]]''<ref name=B87/><ref name=S95/> |
|||
|''Qarunavus meyeri'' |
|||
|Lower Sequence |
|||
|Quarry A |
|||
|Mandibular material of a juvenile. |
|||
|A [[Ptolemaiidae|ptolemaiid]] afrothere. Despite being from a younger animal, the material of ''Q. meyeri'' is larger than that of ''Ptolemaia lyonsi'' which is known from the same locality. Simons and Rasmussen suggest it may have been a racoon-like omnivore.<ref name=K23/> |
|||
| |
|||
|- |
|||
|} |
|||
'''Primates''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Aegyptopithecus]]''<ref name=S13/><ref name=K23/> |
|||
|''Aegyptopithecus zeuxis'' |
|||
|Upper Sequence |
|||
| |
|||
|Mandibular fragments and complete skulls of both males and females. |
|||
|A [[Propliopithecidae|propliopithecid]] primate from the upper sequences of the formation. It may be synonymous with ''[[Propliopithecus]]''. |
|||
|[[File:Aegyptopithecus zeuxis paris.jpg|frameless|center]] |
|||
|- |
|||
|''[[Aframonius]]''<ref name=K23/> |
|||
|''Aframonius dieides'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Three partial mandibles. |
|||
|An adapid primate |
|||
| |
|||
|- |
|||
|''[[Afrotarsius]]''<ref name=S13/> |
|||
|''Afrotarsius chatrathi'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A single mandible. |
|||
|A primate of controversial classification. It reached an estimated bodymass of {{convert|130-300|g|abbr=on}}. |
|||
| |
|||
|- |
|||
|''[[Anchomomys]]''<ref name="Seiffert"/><ref name=K23/> |
|||
|''Anchomomys milleri'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
| |
|||
| |
|||
|- |
|||
|rowspan=3| |
|||
''[[Apidium]]''<ref name=S13/><ref name=K23/> |
|||
|''Apidium bowni'' |
|||
|Upper Sequence |
|||
|Quarry V |
|||
| |
|||
| |
|||
|rowspan=3| |
|||
[[File:Apidium phiomense 03.jpg|frameless|center]] |
|||
|- |
|||
|''Apidium moustafai'' |
|||
|Upper Sequence |
|||
|Quarry G |
|||
| |
|||
| |
|||
|- |
|||
|''Apidium phiomense'' |
|||
|Upper Sequence |
|||
| |
|||
|Various remains including cranial and limb bones. |
|||
|A parapithecid primate known from a rich fossil record. ''Apidium phiomense'' is the type species and one of the first primates described from Jebel Qatrani. |
|||
|- |
|||
|''[[Arsinoea]]''<ref name=S01/><ref name="Simons1992">{{Cite journal | first = E. L. | last = Simons | year = 1992 | title = Diversity in the early Tertiary anthropoidean radiation in Africa | journal = Proceedings of the National Academy of Sciences | volume = 89 | issue = 22 | pages = 10743–10747 | doi=10.1073/pnas.89.22.10743| pmc = 50418 | pmid=1438271| bibcode = 1992PNAS...8910743S | doi-access = free }}</ref> |
|||
|''Arsinoea kallimos'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A mandible with teeth |
|||
|A parapithecid primate. |
|||
| |
|||
|- |
|||
|''[[Catopithecus]]''<ref name=S01/><ref name=S13>{{Cite journal|last1=Seiffert|first1=E. R.|last2=Simons|first2=E. L.|year=2013|title=Last of the oligopithecids? A dwarf species from the youngest primate-bearing level of the Jebel Qatrani Formation, northern Egypt|journal=Journal of Human Evolution|volume=64|issue=3|pages=211–215|doi=10.1016/j.jhevol.2012.10.011}}</ref><ref name=Simons89>{{Cite journal|last1=Simons|first1=E.L.|year=1989|title=Description of two genera and species of late Eocene Anthropoidea from Egypt.|url= https://www.pnas.org/doi/pdf/10.1073/pnas.86.24.9956| journal=Proceedings of the National Academy of Sciences|volume=86|issue=24|pages=9956-9960}}</ref> |
|||
|''Catopithecus browni'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Over 16 specimens including crania, mandibles and postcranial remains. |
|||
|A very common species of [[Oligopithecidae|oligopithecid]]. It was similar in size to large [[Callitrichidae|callitrichid]] primates and smaller [[night monkey]]s, [[squirrel monkeys]] and [[titi monkey]]s. Its teeth suggest a diet of leaves. |
|||
|[[File:Catopithecus browni.jpg|frameless|center]] |
|||
|- |
|||
|''[[Oligopithecus]]''<ref name=S13/> |
|||
|''Oligopithecus savagei'' |
|||
|Lower Sequence |
|||
|Quarry E |
|||
|A partial hemimandible and isolated teeth. |
|||
|A rarer species of oligopithecid. Members of this family are almost absent in the upper sequence of the formation, which may be a delayed result of climate change during the Eocene-Oligocene transition. The teeth of Oligopithecus indicate it may have been a frugivore. |
|||
| |
|||
|- |
|||
|[[Oligopithecidae]] indet.<ref name=S13/> |
|||
| |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A left hemimandible with alveoli for all teeth and a single, mostly complete molar tooth. |
|||
|The smallest known oligopithecid and one of the smallest anthropoids. It is comparable in body mass to modern marmosets of the genus ''[[Callithrix]]'' and has been noted to be smaller than the [[fat-tailed dwarf lemur]]. |
|||
| |
|||
|- |
|||
|''[[Parapithecus]]''<ref name=S13/><ref name=K23/> |
|||
|''Parapithecus fraasi'' |
|||
|Upper Sequence |
|||
| |
|||
|A complete mandible. |
|||
|A parapithecid primate. It has been suggested to by a synonym of ''Apidium'', however this view is not universally accepted. |
|||
| |
|||
|- |
|||
|''[[Plesiopithecus]]''<ref name="Simons1992"/> |
|||
|''Plesiopithecus teras'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A mandible and a nearly complete skull. |
|||
|An enigmatic lemur-like primate belonging to the family [[Plesiopithecidae]]. It may be related to the modern [[aye-aye]]. |
|||
|rowspan=2| |
|||
[[File:Propliopithecus haeckeli mandible.JPG|frameless|center]] |
|||
|- |
|||
|rowspan=2| |
|||
''[[Propliopithecus]]''<ref name=S13/><ref name="Seiffert"/><ref name=K23/> |
|||
|''Propliopithecus ankeli'' |
|||
|Upper Sequence |
|||
|Quarry V |
|||
|Dental material. |
|||
| |
|||
|- |
|||
|''Propliopithecus haeckeli'' |
|||
| |
|||
| |
|||
|Multiple remains. |
|||
|A propliopithecid primate from the upper sequences of the formation. ''[[Moeripithecus]]'' and ''[[Aelopithecus]]'' considered to be a synonym of this taxon. |
|||
| |
|||
|- |
|||
|''[[Proteopithecus]]''<ref name=Simons89/><ref name=K23/> |
|||
|''Proteopithecus sylviae''<ref name=S01/> |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A left maxillary fragment and additional remains including a partial skull. |
|||
|A [[Proteopithecidae|proteopithecid]] primate. |
|||
| |
|||
|- |
|||
|rowspan=3| |
|||
''[[Qatrania]]''<ref name=S01/><ref name=S83>{{Cite journal|last1=Simons|first1=E.L.|last2=Kay|first2=R.F.|year=1983|title= Qatrania, new basal anthropoid primate from the Fayum, Oligocene of Egypt|journal=Nature| volume=304|pages=624–626}}</ref><ref name=K23/><ref name=S13/> |
|||
|''Qatrania basiodontos'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|Mandibular remains |
|||
|Originally described as ''Abuqatrania basiodontos''. |
|||
| |
|||
|- |
|||
|''Qatrania fleaglei'' |
|||
|Upper Sequence |
|||
|Quarry M |
|||
|A fragmentary hemimandible. |
|||
|A parapithecid primate with an estimated bodymass of {{convert|300-500|g|abbr=on}}. |
|||
| |
|||
|- |
|||
|''Qatrania wingi'' |
|||
|Lower Sequence |
|||
|Quarry E |
|||
|Fragmentary dentail remains. |
|||
|A small parapithecid primate weighing less than {{convert|300|g|abbr=on}}. |
|||
| |
|||
|- |
|||
|''[[Serapia]]''<ref name=S01>{{Cite journal|last1=Simons|first1=E. L.|last2=Seiffert|first2=E. R.|last3=Chatrath|first3=P. S.|last4=Attia|first4=Y.|year=2001|title=Earliest Record of a Parapithecid Anthropoid from the Jebel Qatrani Formation, Northern Egypt|journal=Folia Primatologica|volume=72|issue=6|pages=316–331|doi=10.1159/000052748}}</ref><ref name=K23/> |
|||
|''Serapia eocaena'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
|A partial mandible |
|||
|A proteopithecid primate. |
|||
| |
|||
|- |
|||
|''[[Simonsius]]''<ref name=K23/> |
|||
|''Simonsius grangeri'' |
|||
|Upper Sequence |
|||
|Quarry I |
|||
|A mandibular fragment. |
|||
|A disputed genus of proteopithecid primate. It was originally named as a species of ''Parapithecus'', but later raised to its own genus. However, this is not universally accepted. |
|||
|[[File:Parapithecus grangeri mandible 2.jpg|frameless|center]] |
|||
|- |
|||
|''[[Wadilemur]]''<ref name=K23/> |
|||
|''Wadilemur elegans'' |
|||
|Lower Sequence |
|||
|Quarry L-41 |
|||
| |
|||
|A possible [[Lorisiformes|lorisiform]]. |
|||
| |
|||
|- |
|||
|} |
|||
'''Proboscidae''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Moeritherium]]''<ref name=K23/> |
|||
|''Moeritherium trigodon'' |
|||
| |
|||
| |
|||
|A right mandibular ramus and other remains. |
|||
|A small, possibly semi-aquatic proboscidean roughly the size of a tapir. ''M. trigodon'' has some features of its dentition that may suggest that it could be a genus distinct from Moeritherium. Another species was also named from the formation, ''M. andrewsi'', but modern research has drastically cut down on the amount of valid species, with most now being lumped into either the type species of ''M. trigodon''. |
|||
|[[File:Moeritherium andrewsi.jpg|frameless|center]] |
|||
|- |
|||
|rowspan=4| |
|||
[[Palaeomastodon]]<ref name=K23/> |
|||
|''Palaeomastodon beadnelli'' |
|||
| |
|||
| |
|||
|Mandibular remains. |
|||
|An early proboscidean named by Andrews in 1901. It is the type species of the genus ''Palaeomastodon''. Unlike ''Moeritherium'',, ''Palaeomastodon'' and ''Phiomia'' are considered to have been terrestrial animals. |
|||
|rowspan=4| |
|||
[[File:Palaeomastodon beadnelli.jpg|frameless|center]] |
|||
|- |
|||
|''Palaeomastodon minor'' |
|||
| |
|||
| |
|||
| |
|||
|A species distinguished from ''P. beadnelli'' through its smaller size. The status of most ''Palaeomastodon'' species is complex and has been discussed repeatedly. |
|||
|- |
|||
|''Palaeomastodon parvus'' |
|||
| |
|||
| |
|||
| |
|||
|Another small species of ''Palaeomastodon''. |
|||
|- |
|||
|''Palaeomastodon wintoni'' |
|||
| |
|||
| |
|||
| |
|||
|Although morphologically similar to ''P. minor'' it was slightly larger, but fails to approach the size of the type species. It is the most common species in the formation. |
|||
|- |
|||
|''[[Phiomia]]''<ref name=K23/> |
|||
|''Phiomia serridens'' |
|||
| |
|||
| |
|||
|Various remains |
|||
|Although multiple species have been named in the past, ''Phiomia serridens'' is currently the only valid species of this genus native to Fayum. Species of ''Palaeomastodon'' were at times also assigned to ''Phiomia''. |
|||
|[[File:Phiomia serridens skull.JPG|frameless|center]] |
|||
|- |
|||
|} |
|||
'''Sirenia''' |
|||
{| class="wikitable" align="center" width="100%" |
|||
|- |
|||
! Name |
|||
! Species |
|||
! Member |
|||
! Locality |
|||
! Material |
|||
! Notes |
|||
! Image |
|||
|- |
|||
|''[[Eosiren]]''<ref name=Eosiren>{{Cite book|last1=Domning|first1=D.P.|last2=Gingerich|first2=P.D.|last3=Zalmout|first3=I.S.|year=2010|title=Cenozoic Mammals of Africa|chapter=14: Sirenia|pages=147-160}}</ref><ref name=Eos2>{{Cite journal|last1=Domning|first1=D.P.|last2=Gingerich|first2=P.D.|last3=Simons|first3=E.L.|last4=Ankel-Simons|first4=F.A.|year=1994|title=A new early Oligocene dugongid (Mammalia, Sirenia) from Fayum Province, Egypt|url=https://deepblue.lib.umich.edu/bitstream/handle/2027.42/48643/ID510.pdf?sequence=2|journal=Contributions from the Museum of Paleontology The University of Michigan|volume=29|issue=4}}</ref> |
|||
|''Eosiren imenti'' |
|||
|Upper Sequence |
|||
|Quarry O |
|||
|Skull and ribs. |
|||
|''E. imenti'' is the most derived member of the genus ''Eosiren'' and had an elongated head. |
|||
|[[File:Eosiren remains.png|frameless|center]] |
|||
|- |
|||
|} |
|||
== See also == |
== See also == |
||
| Line 55: | Line 1,555: | ||
==References== |
==References== |
||
<references /> |
<references /> |
||
* {{aut|Vickers-Rich, Patricia & Rich, Thomas Hewett}} (1993); ''Wildlife of Gondwana''. Reed. {{ISBN|0-7301-0315-3}} |
|||
{{-}} |
{{-}} |
||
| Line 69: | Line 1,568: | ||
[[Category:Eocene Series of Africa]] |
[[Category:Eocene Series of Africa]] |
||
[[Category:Oligocene Series of Africa]] |
[[Category:Oligocene Series of Africa]] |
||
{{egypt-geo-stub}} |
|||
{{Paleogene-stub}} |
|||
{{Africa-geologic-formation-stub}} |
|||
Revision as of 16:58, 10 May 2023
| Jebel Qatrani Formation | |
|---|---|
| Stratigraphic range: Eocene-Oligocene ~ | |
 | |
| Type | Formation |
| Sub-units | Lower Sequence (Lower Fossil Wood Zone)
Barite Sandstone Upper Sequence (Upper Fossil Wood Zone) |
| Underlies | Widan el Faras Basalt |
| Overlies | Qasr el Sagha Formation |
| Area | Fayum Depression |
| Thickness | 340 m |
| Lithology | |
| Primary | sandstone, mudstone |
| Location | |
| Region | Faiyum Governorate |
| Country | |
The Jebel Qatrani Formation (also Gebel Qatrani, Gabal Qatrani or Djebel Qatrani) is a geologic formation located in the Faiyum Governorate of central Egypt. It is exposed between the Jebel Qatrani escarpment and the Qasr el Sagha escarpment, north of Birket Qarun lake near Faiyum. The formation conformably overlies the Qasr el Sagha Formation and is topped by the Widan el Faras Basalt. The age of the formation has been subject to debate, but the most recent research indicates that it covers both the latest parts of the Eocene and the Early Oligocene, spanning over the boundry between these two time periods.
The geology and fauna of this formation gives a good idea of the environment and animals present during this time period. Research suggests that the Jebel Qatrani Formation featured a mix of subtropical to tropical forest, lowland swamps and marshes, ponds and rivers that would would empty northward into the Tethys Sea. This is supported by the presence of water-dependent fauna including podocnemidid turtles, crocodilians, sea cows, various fish, jacanas, early flamingo-relatives, ospreys, herons and shoebills.
Besides these, the fossil record of the Jebel Qatrani Formation is especially well known for its value to understanding the early evolution of many modern mammal groups. Primates are represented by over a dozen genera, several forms of early elephants have been recovered from the sediments including the terrestrial Phiomia and the semi-aquatic Moeritherium. The fossil rodents of the formation meanwhile are thought to be an important link between the African phiomorphs (dassies, old world porcupines, mole rats and cane rats) and the caviomorphs of South America (capybaras, chinchillas and new world porcupines). Besides these early members of groups that would later rise to prominence, the formation was also home to a variety of unique groups no longer found today or only found in a greatly diminished diversity. This includes the enigmatic, possibly carnivorous ptolemaiids, large hyaenodonts, a vast number of highly diverse hyracoids including species the size of rhinos, anthracotheres and the bizarre embrithopod Arsinoitherium.
Geography and history
)_(17971592490).jpg)
Outcrops of the Jebel Qatrani Formation are present in the northern Fayum Depression southwest of Cairo.[1] The Fayum Depression is an oasis west of the Nile in northern Egypt.[2]
The formations of the Fayum have been studied for a significant amount of time by numerous paleontologists, with research dating back to as early as the 19th century. During this time the region was studied extensively by scientists including but not limited to Charles William Andrews, Henry Fairfield Osborn, René Fourtau and Ernst Stromer. Among the first names for what is now known as the Jebel Qatrani formation was the “Fluvio-Marine Series”, as coined by Hugh J. L. Beadnell. However, despite the bulk of research conducted in the late 19th and early 20th century, the Fayum localities would eventually enter a period of obscurity following the outbreak of World War I which continued throughout the mid 20th century and World War II. Research resumed during the 1960s, following an expedition under Elwyn LaVerne Simons.[2] Previously, the formations of the region had been primarily known for the preservation of mammals, but Birds were also known from few specimens uncovered in the early 20th century. During the 60s, improved collection methods and additional expeditions by the Yale and Duke University gathered much additional material in association with the Egyptian Geological Survey and the General Petroleum Company.[1] Around the 70s, the scope of the expedition was broadened to account for more diverse fields of study, leading to more precise datings of the strata.[2]
Geology and stratigraphy

The formation overlies the Eocene Qasr el Sagha Formation and is overlaid by the Oligocene Widan el Faras Basalt. The formation contains at least two major fossil bearing layers, one in the upper sequence, which is used to refer to the top layers of the formation, and a second in the lower sequence. These two layers have also been called the Upper and Lower Fossil Wood Zones in older publications. Both sequences of the Jebel Qatrani Formation are separated from one another by the so called Barite Sandstone, a layer with a thickness of 4–10 m (13–33 ft).[1][2]
The dating of the Jebel Qatrani Formation has historically been under debate, with some researchs having placed it either entirely within the Eocene or Oligocene and some arguing that it spans both periods. Rassmussen and colleagues for instance argued that the central Barite Sandstones separating both fossil bearing layers mark the exact Eocene-Oligocene Boundry. Part of the reasoning for this connects to the primate fauna of the formation, with propliopithecids and parapithecines only appearing in the upper localities. Other dating methods meanwhile have generally not been applicable. According to Seiffert, the mammal fauna of the formation on its own is too endemic and marine invertebrates are absent, preventing biostratigraphic dating. Radioisotopic dating was conducted on the overlying Widan el Faras Basalt, however the results of this suggested an age of 23.6 million years for its lower units, much younger than prior estimates for the Jebel Qatrani Formation. However, in a 2006 publication Seiffert draws comparisson between the Fayum fauna and the fossil record of the Ashawq Formation in Oman, which notably preserved vertebrates as well as foraminifera useful in dating. Foraminifera biostrategraphy and magnetostratigraphy suggest that the examined localities in Oman date to approximately 31-31.5 (Taqah locality) and 33.7-33.3 Ma (Thaytiniti locality). Seiffert argues that the mammal fauna from Oman most closely resembles those found in the oldest localities of the Lower Sequence of the Jebel Qatrani Formation (L-41) and the oldest localities of the Upper Sequence (quarries G and V). The hyraxes Thyrohyrax and Saghatherium occur in both formations, as does the primate Moeripithecus. More generally, both formations preserve propliopithecids and parapithecine parapithecids as well as oligopithecids. While the former two groups are restricted to the younger Fayum sequence, the latter is the most common primate family in the older sediments. Assuming the traditional interpretation of the Jebel Qatrani Formation, this would mean that many of these taxa would have had to appear 2 to 4 million years earlier in Egypt than in Oman, which is considered to be unlikely by Seiffert. According to them the formation spans approximately 8 million years, with its oldest localities situated in latest Eocene strata. Locality BQ-2 has been estimated to be 37 million years old (early Priabonian), while L-41 falls into an age range of 34.8–33.7 million years old, a timespan that includes the Eocene-Oligocene Boundry. Although an earliest Oligocene age could not be disproven by Seiffert, he argues that a latest Eocene age should be considered more likely based on an unconformity present just above the locality (one also acknowledged as a possible candidate for the EOB by Rassmussen). This means that only the lower 48 m (157 ft) of the formation are Eocene in age, including both the BQ-2 and L-41 localities. The remainder of the Lower Sequence, as well as the entire Upper Sequence, would subsequently fall within the Oligocene. The position of quarries A and B in regards to the boundry is ambiguous, however Quarry E on the other hand is considered without doubt Oligocene (ca. 33 Ma) in age by Seiffert.[3]
Paleoenvironment

The environment of the Jebel Qatrani formation has been described as a subtropical to tropical lowland plain by Bown, who further suggests the presence of streams and ponds. Based on the fossil bird remains, which includes the fossils of a variety of animals highly associated with water (ospreys, early flamingos, jacanas, herons, storks, cormorants and shoebills), Rasmussen and colleages inferred that the environment featured slow-moving freshwater with a substantial amount of aquatic vegetation, which matches the prior hypothesis. Although lithology suggests that most fossils were deposited on sandbanks after being transported by currents, the authors argue that swamps could have easily formed along the banks of the river that was present during the Oligocene and may account for the mudstone found in certain quarries. They furthermore suggest that the fossil birds of Fayum, due to their affinities with modern groups, should be considered a more valuable indicator of the environment when compared with the fossil mammals, many of which belonged to families lacking modern examples. The absence of other birds typical for such an environment may be explained either through sampling bias or due to the fact that said groups had simply not yet been present in Oligocene Africa. Generally, Rasmussen and colleagues compare the environment of Jebel Qatrani to freshwater habitats in modern Central Africa.[1] The discovery of snakehead fossils seem to support Rasmussen's interpretation, as the genus Parachanna today prefers slow-moving backwaters with plenty of vegetation. Other fish present meanwhile, notably Tylochromis, suggest that deep, open water was likewise present. The river channels may have been overgrown with reeds, papyrus and featured floating vegetation like water lilies and Salvinia.[4] In a 2001 paper Rasmussen et al. argued that the sandstone and mudstone of the formation likely formed as sediments were aggraded by a system of river channels that emptied towards the west into the Tethys. Here they reconstructed the environment as a tropical lowland swamp forest intermingled with marshes. They furthermore suggest that the environment would have experienced monsoons.[5] Overall this indicates that this region was a part of an extensive belt of tropical forest that stretched across what is now northern Africa, which would gradually give rise to open woodland and even steppe the further one was to travel inland.[2]
Paleobiota
Fish
Actinopterygii
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Alestidae indet.[4] | Lower and Upper Sequence | Quarry E, I, M, R & X | Various teeth | Different teeth belonging to Characiformes, they are common across both fossil bearing members. A large range of tooth morphotypes is present in the formation, however this may not reflect the actual diversity of african tetra species present. | ||
| Amiiformes indet.[4] | ||||||
| Lates[4] | Lates sp. | Lower and Upper Sequence | Quarry E, I, M & V | Two crania and various other isolated material. | Its absence from the lacustrine L-41 environment might indicate that the water was too shallow. Modern Lates can grow large and prefer large lakes and rivers. | |
| Parachanna[4] | Parachanna fayumensis | Lower and Upper Sequence | Quarry L-14, P & M | Various cranial remains including a dentary. | A snakehead fish, the Jebel Qatrani material may suggest that this group arrived in Africa earlier than it had been previously thought. | |
| Siluriformes indet.[4] | Lower and Upper Sequence | Quarry L-41, B, G, E, I, J, K, M, P & V | Three skulls and various postcranial remains, predominantly fin spines. | The three known skulls are approximately the same size, but may not represent the same taxon. The postcranial elements indicate a greater size range. In some aspects the Jebel Qatrani catfish resemble the genus Fajumia. | ||
| cf. Tylochromis[4] | Lower and Upper Sequence | Quarry E, I & X | A lower jaw and additional material including teeth. | A basal cichlid. |
Chondrichthyes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Dasyatoidea indet.[4] | ||||||
| Lamniformes indet.[4] |
Sarcopterygii
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Dipnoi indet.[4] | Quarry L-41 |
Reptiles
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Albertwoodemys[6] | Albertwoodemys testudinum | Lower Sequence | A locality west of the AMNH quarries. | A partial plastron with articulated peripheral elements of the upper shell. | A side-necked turtle of the family Podocnemididae with a high-domed shell likely similar to tortoises. It may be a related taxon to specimen UCMP 42008, from the Miocene of Kenya. | |
| Andrewsemys[6][7] | Andrewsemys libyca | North of Lake Qarun | Various shell remains. | A side-necked turtle of the family Podocnemididae previously known as "Stereogenys" lybica. The holotype specimen, among the best preserved turtle shells from Fayum, was thought to be lost before being rediscovered and used to errect a new genus. Several additional specimens are known from the Jebel Qatrani and its underlying formations. | ||
| "Crocodylus" megarhinus[8] | Various cranial and mandibular remains | A large and broad-snouted species of crocodilian incorrectly assigned to Crocodylus. A second Fayum crocodile, "Crocodylus" articeps, has been considered a younger individual of this species by Christopher Brochu. | ||||
| Dacquemys[6][9] | Dacquemys paleomorpha | Lower Sequence | Quarry B | A nearly complete skull | A side-necked turtle of the family Podocnemididae that may be a related taxon to specimen UCMP 42008. It is possible that Dacquemys represents skull material of Albertwoodemys. | |
| Eogavialis[8] | Eogavialis africanum | An early genus of gavialoid common in the formation. Two species have originally been named from Jebel Qatrani,Eogavialis gavialoides and Eogavialis tenuirostre (both originally as Tomistoma), however recent research suggests that all Fayum Eogavialis species including those of other formations in the region may simply represent a single species.[2] | ||||
| Gigantochersina[10] | Gigantochersina ammon | Lower Sequence | Quarry A & B | Multiple specimens including a nearly complete carapace and plastron as well as a partial pelvis | The oldest known tortoise from Africa. Testudo beadnelli and Testudo isis were both named, but have since then been synonymized with G. ammon.[2] | |
| Neochelys[6] | Neochelys fajumensis | Lower and Upper Sequence | Quarry L-41, A, B, C, I, M, O, P & R | Various remains of the plastron and carapace as well as skull material. | A side-necked turtle of the family Podocnemididae, similar to Erymnochelys and previously assigned to said genus. The type specimen, an anterior plastron, has potentially been lost. They are among the most common fossils in the quarries L-41, I and M. | |
| Varanidae indet.[2] | Upper Sequence | Quarry I | A presacral vertebra | I single bone described by Smith et al. and originally assigned to the genus Varanus. Later research by Holmes et al. suggests it was a stem-varanid, distinct from other remains found in the formation. | ||
| Varanus[2] | Upper Sequence | Quarry I & M | Vertebrae | Isolated bones described by Holmes et al. and considered to be part of the modern genus Varanus. They argue that this represents the oldest known member of the genus and indicates an African origin for modern monitor lizards. |
Birds
Accipitriformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Accipitridae indet., aff. Haliaeetus[1] | Lower Sequence | Quarry A | The distal end of a tarsometatarsus. | A bird of prey similar in size and morphology to the modern sea eagles. The early strata of the formation indicate closer proximity to the shore, possibly supporting a lifestyle similar to modern Haliaeetus species. | ||
| Pandionidae indet., aff. Pandion[1] | Upper Sequence | Quarry M | The distal end of a humerus. | A fossil nearly identical to the extant osprey, but smaller. | ||
| Pandionidae? indet.[1] | Upper Sequence | Quarry I | A damaged carpometacarpus. | A raptorial bird approximately the size of the extant osprey, however more robust. |
Charadriiformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Janipes[1] | Janipes nymphaeobates | Upper Sequence | Quarry M | The distal end of a tarsometatarsus | A large species of jacana, exceeding all modern taxa in size but still smaller than Nupharanassa bulotorum. The Fayum jacanas already show signs of a lifestyle like that of their modern relatives and subsequently may indicate a dense floating vegetation. | |
| Nupharanassa bulotorum | Upper Sequence | Quarry M | Multiple specimens preserving the tarsometatarsus. | A large species of jacana, between 30-35% larger than the largest extant species, the bronze-winged jacana. | ||
| Nupharanassa tolutaria | Lower Sequence | Quarry E | The distal end of a tarsometatarsus. | A smaller relative of Nupharanassa bulotorum, it's distinguished by its much smaller size and older age. It is the only known jacana in the lower sequence of the Jebel Qatrani Formation. |
Ciconiiformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Palaeoephippiorhynchus[1] | Palaeoephippiorhynchus dietrichi | Upper Sequence | Quarry M | A nearly complete rostrum with mandible and partial cranium and the distal end of a right tibiotarsus. | A large stork comparable in size to the modern jabiru and marabou stork. While the type material bears great resemblance to the extant saddlebill, a tibiotarsus found in a different quarry shares no similarities with any modern taxa and may or may not belong to Palaeoephippiorhynchus. |
Cuculiformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Musophagidae indet., aff. Crinifer[1] | Upper Sequence | Quarry M | The distal ends of a tarsometatarsus and a humerus. | A relatively large type of turaco and among the earliest recorded members of the group. It shares similarities to the extant genus Crinifer. |
Gruiformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Gruidae indet.[1] | Upper Sequence | Quarry M | The distal end and partial shaft of a tarsometatarsus. | A bird bearing resemblance to modern crowned cranes, cranes of the genus Grus and the limpkin. It's size is between that of the limpkin and the demoiselle crane, although closer to the former. | ||
| Rallidae indet.[1] | Upper Sequence | Quarry M | The distal end of a tarsometatarsus. | Possibly a type of rail bearing resemblance to the genera Sarothrura, Coturnicops and Laterallus. |
Paleognathae
| Name | Species | Member | Material | Notes | Image |
|---|---|---|---|---|---|
| Eremopezus[1][5] | Eremopezus eocaenus | Lower Sequence | Various fragmentary leg bones including the distal end of a tibiotarsus and the distal end of a tarsometatarsus. | An enigmatic large groundbird comparable in size to a modern rhea. It was initially thought to be a type of ratite, but its relationship with other birds has repeatedly been questioned. Stromeria fajumensis, found in the same formation, is thought to be synonymous with Eremopezus. |
Pelecaniformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Ardeidae indet.[1] | Upper Sequence | Quarry I & M | A partial rostrum, a distal tarsometatarsus of a juvenile specimen and the first phalanx of the third toe. | A medium sized heron comparable in size to the reddish egret and the great white egret. The lack of overlapping material means that it is uncertain if these fossils represent one or several species. | ||
| Goliathia[1] | Goliathia andrewsi | Lower and Upper Sequence | Quarry M | A complete ulna and the distal end of a tarsometatarsus. | A large relative of the modern shoebill. |  |
| Nycticorax[1] | Nycticorax sp. | Upper Sequence | Quarry M | A nearly complete tarsometatarsus and a coracoid. | A bird who's remains are described as being identical to that of the extant black-crowned night heron, indicating that night herons diverged from other herons at least 31 million years ago. | |
| Xenerodiops[1] | Xenerodiops mycter | Upper Sequence | Quarry I & M | A nearly complete rostrum and a humerus lacking its distal end. | An unusual heron with a heavy and downwards curved beak. |
Phoenicopteriformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Phoenicopteridae indet. Species 1, aff. Palaelodus[1] | Upper Sequence | Quarry M & I | Multiple specimens including the tibiotarsus and parts of the metacarpals. | Several bones of the Fayum phoenicopteriform show both similarities to derived flamingos as well as to the more basal Palaeolodus, which Rasmussen and colleagues considered Phoenicopterids rather than placing it within its own family. The material suggests a bird approximately the size of Palaelodus gracilipes. | ||
| Phoenicopteridae indet. Species 2[1] | Upper Sequence | Quarry M | A coracoid fragment | A second, larger phoenicopteriform, closer in size to Palaeolodus crassipes and the lesser flamingo. |
Suliformes
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Phalacrocoracidae indet.[1] | Upper Sequence | Quarry M | A distal premaxilla. | An indetermined cormorant showing a more gradually tapering and more strongly hooked beak. Among modern cormorants, it most closely resembles the Guanay cormorant. |
Mammals
Afroinsectivora
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Eochrysochloris[11] | Eochrysochloris tribosphenus | Lower Sequence | Quarry E | A tiny member of the Afrosoricida. | ||
| Herodotius[12] | Herodotius pattersoni | Lower Sequence | Quarry L-41 | Fossils of the dentary and maxilla. | A species of elephant shrew. | |
| Jawharia[13][11] | Jawharia tenrecoides | Lower Sequence | Quarry E | A relative of tenrecs and golden moles. | ||
| Metoldobotes[2] | Metoldobotes stromeri | A genus of elephant shrew. | ||||
| Qatranilestes[13][11] | Qatranilestes oligocaenus | Upper Sequence | Quarry I | A badly preserved mandible. | The most derived member of the Afrosoricida in the formation. | |
| Widanelfarasia bowni | Lower Sequence | Quarry L-41 | A stem-afrosoricid. | |||
| Widanelfarasia rasmusseni | Lower Sequence | Quarry L-41 |
Artiodactyla
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Bothriogenys andrewsi | Upper Sequence | L-75 | Teeth and dentary remains. | A large anthracothere from the uppermost levels of the formation. |
 | |
| Bothriogenys fraasi | Upper Sequence | Quarry I | A species of Bothriogenys arising from B. gorringei. It may have either split into two lineages or given rise to B. andrewsi through anagenesis. | |||
| Bothriogenys gorringei | Lower Sequence | Quarry M | Originally described based on a mandible. | The oldest species of Bothriogenys in Jebel Qatrani, it may have split into two lineages giving rise to B. fraasi and B. rugulosus. It may have been a semi-aquatic browser or grazer, feeding primarily on foliage. | ||
| Bothriogenys rugulosus | Upper Sequence | A species of Bothriogenys that likely evolved from B. gorringei. | ||||
| Nabotherium[16][18][2] | Nabotherium aegyptiacum | A partial skull, several fossils of the mandible and maxilla as well as isolated teeth. | An anthracothere with well developed canine teeth. Its dentition is better suited to crushing fruit rather than slicing vegetation. | |||
| Qatraniodon[14][16][19] | Qatraniodon parvus | Lower Sequence | The holotype consists of a partial mandible. | The smallest anthracothere from the Jebel Qatrani Formation and not as common than Bothriogenys. |
Chiroptera
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Dhofarella[20][21] | Dhofarella sigei | Lower Sequence | Quarry L-41 | A left dentary. | A type of sheath-tailed bat. | |
| Khonsunycteris[20] | Khonsunycteris aegypticus | Lower Sequence | Quarry L-41 | A left dentary. | A type of vesper bat. | |
| Phasmatonycteris[22][20] | Phasmatonycteris phiomensis | Upper Sequence | Quarry I | Dentary remains. | A type of sucker-footed bat. | |
| Philisis[20][21] | Philisis sphingis | Upper Sequence | Quarry I | Material of the maxilla and dentary. | A philisid bat related to the older Witwatia from Birket Qarun. | |
| Saharaderma[20] | Saharaderma pseudovampyrus | Lower Sequence | Quarry L-41 | A right dentary. | A type of false vampire bat. | |
| Vampyravus[20][21] | Vampyravus orientalis | Upper Sequence | A large humerus. | A bat of uncertain affinities. It was the first fossil bat discovered in Africa and the largest bat of the Fayum succession, weighing up to 120 g (4.2 oz). This puts it within the size range of the Egyptian fruit bat. |
Embrithopoda
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Arsinoitherium[23][24] | Arsinoitherium zitteli | Lower and Upper Sequence | Just above Quarry L-41 to Quarry M, spanning most of the sequence. | A large arsinoitheriid embrithopod well known for its two horns. A large amount of material is known and in the past a second species from Jebel Qatrani, A. andrewsi, has been proposed. Some researchers however consider it to be a synonym of A. zitteli with the differences possibly being caused by sexual dimorphism. |  |
Hyaenodonta
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Akhnatenavus leptognathus | Lower Sequence | Quarry A | A slender mandible. | A member of the Hyainailourinae, it was originally described as Pterodon leptognathus. It was smaller than the two other "Pterodon" species from Jebel Qatrani and Metapterodon. |  | |
| Akhnatenavus nefertiticyon | Lower Sequence | Quarry L-41 | A complete cranium as well as a palate and dentary. | A second species of Akhnatenavus smaller than A. leptognathus. The skull also appears to have been much shorter than in the type species. Calculations resulted in an average bodymass of 19.2 kg (42 lb), approximately as large as a Eurasian lynx or wolverine. |  | |
| Apterodon altidens | Lower Sequence | Described based on a maxilla. | Although named in 1910, A. altidens was not properly described until 1911. A mandible was also referred to this species, but this decision has been considered dubious by Lange-Badré and Böhme. | |||
| Apterodon macrognathus | Lower Sequence | Quarry A & B | Skulls and mandibles | A hyaenodont originally described as a species of Pterodon. |  | |
| Brychotherium[25][30] | Brychotherium ephalmos | Lower Sequence | Quarry L-41 | Multiple lower jaws and parts of the rostrum. | A teratodontine hyaenodont approximately as heavy as a modern red fox or American badger. Although the name was coined in a dissertation in 1994, it was not formally described until 2016. |  |
| Falcatodon[31][32][27] | Falcatodon schlosseri | Upper Sequence | Quarry V | Described based on a left dentary. | Erected by Holroyd as a species of Metapterodon, it was even then noted to be possibly distinct. Morales and Pickford later raised it to a distinct genus. There may be additional material from Quarry B in the lower sequence, however the assignment of said fossils is only tentative. |  |
| Masrasector aegypticum | Upper Sequence | Quarry G | Dental remains | A teratodontine hyaenodont smaller than Brychotherium. | ||
| Masrasector nananubis | Lower Sequence | Quarry L-41 | Skulls, mandibles and multiple humeri. | A smaller species of Masrasector from older deposits. It's estimated boy size was within the range of modern striped skunks and small-spotted genets. The limb bones indicate it was a fast runner that could have hunted the local rodents. | 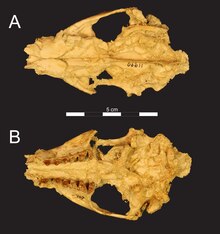 | |
| Metapterodon[26][27][15] | Metapterodon brachycephalus | Lower Sequence | Quarry A | A left dentary. | Named as a species of Hyaenodon, it was eventually placed in Metapterodon by Holroyd. While Morales and Pickford restrict the genus to the Miocene and place Holroyd's other species in separate genera, they do not mention M. brachycephalus. |  |
| Metasinopa[26][25][30] | Metasinopa fraasi | Upper Sequence | A nearly complete lower jaw. | A possible relative to teratodontines, although its exact relationship with other hyaenodontids needs more testing. It was larger than Masrasector. |  | |
| "Pterodon" africanus | Lower Sequence | Quarry A | Mandibles, a rostrum and an assigned neck vertebra. | The first hyaenodont described from Fayum, later analysis showed that it was much more derived than Pterodon dasyuroides, the type species. Consequently, a different genus name is required. |  | |
| "Pterodon" phiomensis | Lower Sequence | Quarry A | A mandible smaller than that of "P." africanus. | Like "P." africanus, "P." phiomensis is much more derived than the type species of Pterodon and thus requires a different genus name. Furthermore, analysis showed that it was not closely related to "P." africanum either. It is most closely allied with the clade formed by Akhnatenavus, Isohyaenodon and Hyainailouros. |  | |
| "Pterodon" sp. | Upper Sequence | Quarry E | A broken canine. | An isolated tooth similar in size to "P." phiomensis. | ||
| "Pterodon" syrtos | Upper Sequence | Quarry M | A right maxilla fragment. | |||
| Quasiapterodon[29][2][31][33] | Quasiapterodon minutus | Upper Sequence | Originally named as a species of Apterodon alongside A. altidens, later research showed it clearly differed from this genus. It may instead be related to "Sinope" ethiopica. | |||
| Sectisodon[31][32][27] | Sectisodon markgrafi | A left maxilla. | S. markgrafi was initially named by Holroyd as a species of Metapterodon in 1999, but placed in the new genus Sectisodon by Morales and Pickford in 2017. | |||
| "Sinopa"[34][25][29][35] | "Sinopa" ethiopica | A mandibular ramus. | A hyaenodont originally described as a species of Sinopa, although later research clearly shows that it does not belong to the American-Asian genus. It's exact relation to other hyaenodonts remains uncertain until further study, however it has been suggested to be a relative of Quasiapterodon. |
Holroyd also identified several indetermined pterodontine hyaenodonts from various sequences of the Jebel Qatrani Formation, but doesn't identify them beyond subfamily level.[27]
Hyracoidea
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Antilohyrax[36][37] | Antilohyrax pectidens | Lower Sequence | Quarry L-41 | Mandibles, a crushed cranium as well as various incomplete skull fossils and postcranial elements. | A gazelle-like titanohyracine described in 2000. The teeth and limb bones indicate that it was a cursorial browser, similar to modern antelopes and gazelles. | |
| Bunohyrax fajumensis[38][39][40] | Lower and Upper Sequence | Described based on a mandible. | Bunohyrax species were medium-sized hyracoids somewhat resembling pigs in their dentition. They may have been related to Pachyhyrax. | |||
| Bunohyrax major[38][40] | Upper Sequence | Described based on three premolars. | Originally named Geniohyus major, B. major is larger than B. fajumensis. Additionally, its teeth are more bunodont. | |||
| Bunohyrax sp.[36] | Lower Sequence | Quarry L-41 | ||||
| Geniohyus diphycus[41] | Lower Sequence | A medium-sized species. Geniohyus is among the rarer hyracoids of Jebel Qatrani. | ||||
| Geniohyus magnus[40][41] | Lower and Upper Sequence | Quarry V | Various skull remains including maxillary fragments and dentary remains. | It is the smallest species of Geniohyus, with the name originating when it was still considered apart of Saghatherium. It's dentition forms an intermediate between other Geniohyus and Bunohyrax. | ||
| Geniohyus mirus[38][40][41] | Lower Sequence | Described based on a mandible. | The largest of the three named Fayum Geniohyus species and the type species. | |||
| Geniohyus sp.[36] | Lower Sequence | Quarry L-41 | Geniohyus takes the species name from their morphological similarities to pigs, with early research even classifying them as suids. | |||
| Megalohyrax eocaenus[36][38][39][40] | Upper Sequence | Described based on premaxillary and maxillary material. Also includes mandibular remains. | Megalohyrax show strong sexual dimorphism in regards to their body size. It was a common genus in formation. | |||
| Megalohyrax sp.[36][38] | Lower Sequence | Quarry L-41 | An unnamed species of Megalohyrax preserving an interior mandibular fenestra unlike the younger M. eocaenus. | |||
| Pachyhyrax[36][38][39][41] | Pachyhyrax crassidentatus | Upper Sequence | Quarry I, L-46 & M | Skulls, mandibles and teeth including the complete but crushed skull of a young individual. | A hyracoid resembling the contemporary anthracothere Bothriogenys. It is possible that it was a semi-aquatic herbivore. | |
| Saghatherium antiquum[38][43][40][41] | Lower Sequence | Described based on a maxilla | Saghatherium had heavy jaws and possibly fed on nuts and seeds. Furthermore, members of this genus display pronounced sexual dimorphism and may have had complex mating rituals. The species S. sobrina was shown to simply represent female individuals of this speices. |
 | ||
| Sagahtherium humarum[41] | Upper Sequence | Quarry V | Dentaries and teeth. | A small Saghatherium species that differs due to the presence of an internal chamber in the mandible. | ||
| Saghatherium bowni[36][38][43] | Lower Sequence | Quarry L-41 | A common species of Saghatherium named in 1991. | |||
| Selenohyrax[36][38][41] | Selenohyrax chatrathi | Upper Sequence | Quarry V | Several mandibles. | A hyracoid named for its selenodont teeth (molars with crescent-shaped ridges), which are much more developed than in Titanohyrax. These teeth indicate a diet that required it to slice vegetation rather than grind it. It is a small hyrax, only slightly larger than Saghatherium which it might have descendet from. | |
| Thyrohyrax domorictus[38][39] | Upper Sequence | Quarry I, G, L-46, M, P, R, V & X | Primarily mandibular remains and teeth, but also some limb elements and vertebrae. | Thyrohyrax is among the smaller hyracoids of Fayum and resembles today's arboreal Dendrohyrax. Species of Thyrohyrax display little size-related sexual dimorphism, however males do possess enlarged, tusk-like second incisors and chambered mandibles. | ||
| Thyrohyrax litholagus[36][43] | Lower Sequence | Quarry L-41 | A relative of the younger Thyrohyrax pygmaeus, it was somewhat larger than T. meyeri. | |||
| Thyrohyrax meyeri[36][38][43] | Lower Sequence | Quarry L-41 | T. meyeri was very similar to the younger T. domorictus and may have been a close relative. | |||
| Thyrohyrax pygmaeus[38][41] | Lower Sequence | Quarry A-1 | The front of a skull with an associated mandible as well as several teeth from another specimen. | A close relative of direct descendent of T. litholagus. It was originally assigned to Saghatherium magnum, then described as a species of Megalohyrax before being moved into Pachyhyrax and eventually Thyrohyrax. | ||
| Titanohyrax andrewsi[36][40][41] | Lower Sequence | Five specimen including two mandibles and a single maxilla. | Titanohyrax species are the largest hyracoids of the Fayum Depression and form the clade Titanohyracinae with Antilohyrax. Like their smaller relative, they are thought to have been folivores. The lower incisors of T. andrewsi were shaped like spatulas, rather than tusks. | |||
| Titanohyrax angustidens[36][40][41] | Upper Sequence | Quarry V, R & I | Eight specimen including maxillary, premaxillary and mandibular remains. | T. angustidens has the most complex history among Titanohyrax species, initially assigned to Megalohyrax and given the name M. palaeotheroides, it was later recognized as Titanohyrax and variably considered distinct or synonymous with T. andrewsi. The name T. palaeotheroides was not retained as it was a nomen nudum. | ||
| Titanohyrax ultimus[36][40] | Upper Sequence | Four heavily worn teeth. | The largest species of Titanohyrax, weight estimates for T. ultimus range from 600–1,300 kg (1,300–2,900 lb), making it possibly as large as the modern sumatran rhino. | |||
| Titanohyrax sp.[36] | Lower Sequence | Quarry L-41 | An unnamed species from the L-41 Quarry. |
Marsupalia
| Name | Species | Member | Material | Notes | Image | |
|---|---|---|---|---|---|---|
| Peratherium[2][44] | Peratherium africanum | Upper Sequence | Quarry M | Several specimen including a maxilla and three mandibles | A primitive marsupial and the basalmost mammal found in the sediments of the formation. It was at one point known under the name Qatranitherium, however this name has since then been deemed a junior synonym. |
Rodentia
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Acritophiomys adaios | Lower Sequence | Quarry A, B & E | This species also includes several specimen previously assigned to Phiomys andrewsi. However, as it was not adequately described it is considered a nomen nudum. | |||
| Acritophiomys bowni | Lower Sequence | Quarry L-41 | Complete upper and lower dentition as well as parts of the lower jaw and the skull. | A basal member of Phiomorpha and the largest member of the family "Phiomyidae", although the group is now considered to be paraphyletic. It is the only species of Acritophiomys that had been validly named, as both A. adaios and A. woodi were only described in a thesis. | ||
| Acritophiomys woodi | Lower Sequence | Quarry L-41 | The first phiomydid described from the Quarry L-41. However, as it was not adequately described it is considered a nomen nudum. | |||
| Birkamys[45][2][47] | Birkamys korai | Lower Sequence | Quarry L-41 | Cranial and mandibular remains with teeth, including a partially preserved rostrum. | A close relative of Mubhammys. | |
| Gaudeamus aegyptius | Upper Sequence | Quarry E | Mandibles and teeth | The type species of the genus, described by Wood in the 1960s. It is intermediate in size among the Fayum rodents, larger than the small Phiocricetomys but smaller than Metaphiomys. Its phylogeny is enigmatic and it has variably been placed as a sister to old world porcupines or within Caviomorpha. | ||
| Gaudeamus aslius | Lower Sequence | Quarry L-41 | A distorted skull and various fragmentary remains. | It shows several primitive features and may resemble the ancestral form of this genus. | ||
| Gaudeamus hylaeus | Lower Sequence | Quarry L-41 | A flattened skull and various additional fragments. | G. hylaeus was initially named by Holroyd in a PhD Thesis, rendering it a nomen nudum until the full description authored by Sallam, Seiffert and Simons. | ||
| Metaphiomys beadnelli | Upper Sequence | A left mandible and additional material. | A phiomyid rodent. | |||
| Metaphiomys schaubi | A second species of Metaphiomys described by Wood in the 1960s. | |||||
| Monamys[46][2][49] | Monamys simonsi | Upper Sequence | Quarry I & M | Mandibular remains and teeth. | Previously known as Paraphiomys simonsi, later research has shown that this genus is distinct from the much later Miocene Paraphiomys. | |
| Mubhammys[2][47] | Mubhammys vadumensis | Lower Sequence | Quarry L-41 | Cranial and mandibular elements with teeth. | A close relative of Birkamys. | |
| Phiocricetomys[45][46] | Phiocricetomys minutus | Upper Sequence | Quarry I | A mandible preserving dentition from the 4th deciduous premolar to 2nd molar. | The youngest known member of Phiocricetomyinae. | |
| Phiomys andrewsi | Lower Sequence | Quarry A & B | Described based on partial mandibles. | A phiomyid rodent. | ||
| Phiomys paraphiomyoides | A phiomyid described by Wood in the 1960s. | |||||
| Qatranimys[45] | Qatranimys safroutus | Lower Sequence | Quarry L-41 | Skull remains and teeth. | A diminutive phiocricetomyine rodent. | |
| Talahphiomys[45][46][50] | Talahphiomys lavocati | Lower Sequence | Quarry E | Deciduous premolars and molars. | Initially thought to be a species of Phiomys, it was named Elwynomys in a doctoral dissertation from 1994. Due to the rules of the ICZN, the name Talahphiomys later took precedence. |
Pholidota
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Pholidota[2] | Upper Sequence | Quarry M & L-12 | Two finger bones. | The fragmentary remains of what may have been a pangolin. The presence of termite nests bearing excavation marks may support this, however the identity of these finger bones is not universally supported and has been questioned. |
Ptolemaiida
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Cleopatrodon ayeshae | Upper Sequence | Quarry V | Mandibular remains and teeth. | A ptolemaiid afrothere. The type species of Cleopatrodon. | ||
| Cleopatrodon robusta | Upper Sequence | Quarry I | Mandibular remains and teeth. | C. robusta had more robust jaws than C. ayeshae. | ||
| Ptolemaia grangeri[51] | Upper Sequence | Quarry V | Multiple teeth and a mandibular ramus as well as an almost complete but distorted skull. | A carnivorous afrotherian mammal of the family Ptolemaiidae. It is larger than the older P. lyonsi. | ||
| Ptolemaia lyonsi[53] | Lower Sequence | Quarry A | Lower jaw material. | A carnivorous afrothere first described by Osborn in 1908. | .jpg) | |
| Qarunavus[51][52] | Qarunavus meyeri | Lower Sequence | Quarry A | Mandibular material of a juvenile. | A ptolemaiid afrothere. Despite being from a younger animal, the material of Q. meyeri is larger than that of Ptolemaia lyonsi which is known from the same locality. Simons and Rasmussen suggest it may have been a racoon-like omnivore.[2] |
Primates
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Aegyptopithecus[54][2] | Aegyptopithecus zeuxis | Upper Sequence | Mandibular fragments and complete skulls of both males and females. | A propliopithecid primate from the upper sequences of the formation. It may be synonymous with Propliopithecus. |  | |
| Aframonius[2] | Aframonius dieides | Lower Sequence | Quarry L-41 | Three partial mandibles. | An adapid primate | |
| Afrotarsius[54] | Afrotarsius chatrathi | Upper Sequence | Quarry M | A single mandible. | A primate of controversial classification. It reached an estimated bodymass of 130–300 g (4.6–10.6 oz). | |
| Anchomomys[3][2] | Anchomomys milleri | Lower Sequence | Quarry L-41 | |||
| Apidium bowni | Upper Sequence | Quarry V |
 | |||
| Apidium moustafai | Upper Sequence | Quarry G | ||||
| Apidium phiomense | Upper Sequence | Various remains including cranial and limb bones. | A parapithecid primate known from a rich fossil record. Apidium phiomense is the type species and one of the first primates described from Jebel Qatrani. | |||
| Arsinoea[55][56] | Arsinoea kallimos | Lower Sequence | Quarry L-41 | A mandible with teeth | A parapithecid primate. | |
| Catopithecus[55][54][57] | Catopithecus browni | Lower Sequence | Quarry L-41 | Over 16 specimens including crania, mandibles and postcranial remains. | A very common species of oligopithecid. It was similar in size to large callitrichid primates and smaller night monkeys, squirrel monkeys and titi monkeys. Its teeth suggest a diet of leaves. |  |
| Oligopithecus[54] | Oligopithecus savagei | Lower Sequence | Quarry E | A partial hemimandible and isolated teeth. | A rarer species of oligopithecid. Members of this family are almost absent in the upper sequence of the formation, which may be a delayed result of climate change during the Eocene-Oligocene transition. The teeth of Oligopithecus indicate it may have been a frugivore. | |
| Oligopithecidae indet.[54] | Upper Sequence | Quarry M | A left hemimandible with alveoli for all teeth and a single, mostly complete molar tooth. | The smallest known oligopithecid and one of the smallest anthropoids. It is comparable in body mass to modern marmosets of the genus Callithrix and has been noted to be smaller than the fat-tailed dwarf lemur. | ||
| Parapithecus[54][2] | Parapithecus fraasi | Upper Sequence | A complete mandible. | A parapithecid primate. It has been suggested to by a synonym of Apidium, however this view is not universally accepted. | ||
| Plesiopithecus[56] | Plesiopithecus teras | Lower Sequence | Quarry L-41 | A mandible and a nearly complete skull. | An enigmatic lemur-like primate belonging to the family Plesiopithecidae. It may be related to the modern aye-aye. |
 |
| Propliopithecus ankeli | Upper Sequence | Quarry V | Dental material. | |||
| Propliopithecus haeckeli | Multiple remains. | A propliopithecid primate from the upper sequences of the formation. Moeripithecus and Aelopithecus considered to be a synonym of this taxon. | ||||
| Proteopithecus[57][2] | Proteopithecus sylviae[55] | Lower Sequence | Quarry L-41 | A left maxillary fragment and additional remains including a partial skull. | A proteopithecid primate. | |
| Qatrania basiodontos | Lower Sequence | Quarry L-41 | Mandibular remains | Originally described as Abuqatrania basiodontos. | ||
| Qatrania fleaglei | Upper Sequence | Quarry M | A fragmentary hemimandible. | A parapithecid primate with an estimated bodymass of 300–500 g (11–18 oz). | ||
| Qatrania wingi | Lower Sequence | Quarry E | Fragmentary dentail remains. | A small parapithecid primate weighing less than 300 g (11 oz). | ||
| Serapia[55][2] | Serapia eocaena | Lower Sequence | Quarry L-41 | A partial mandible | A proteopithecid primate. | |
| Simonsius[2] | Simonsius grangeri | Upper Sequence | Quarry I | A mandibular fragment. | A disputed genus of proteopithecid primate. It was originally named as a species of Parapithecus, but later raised to its own genus. However, this is not universally accepted. |  |
| Wadilemur[2] | Wadilemur elegans | Lower Sequence | Quarry L-41 | A possible lorisiform. |
Proboscidae
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Moeritherium[2] | Moeritherium trigodon | A right mandibular ramus and other remains. | A small, possibly semi-aquatic proboscidean roughly the size of a tapir. M. trigodon has some features of its dentition that may suggest that it could be a genus distinct from Moeritherium. Another species was also named from the formation, M. andrewsi, but modern research has drastically cut down on the amount of valid species, with most now being lumped into either the type species of M. trigodon. |  | ||
| Palaeomastodon beadnelli | Mandibular remains. | An early proboscidean named by Andrews in 1901. It is the type species of the genus Palaeomastodon. Unlike Moeritherium,, Palaeomastodon and Phiomia are considered to have been terrestrial animals. |
 | |||
| Palaeomastodon minor | A species distinguished from P. beadnelli through its smaller size. The status of most Palaeomastodon species is complex and has been discussed repeatedly. | |||||
| Palaeomastodon parvus | Another small species of Palaeomastodon. | |||||
| Palaeomastodon wintoni | Although morphologically similar to P. minor it was slightly larger, but fails to approach the size of the type species. It is the most common species in the formation. | |||||
| Phiomia[2] | Phiomia serridens | Various remains | Although multiple species have been named in the past, Phiomia serridens is currently the only valid species of this genus native to Fayum. Species of Palaeomastodon were at times also assigned to Phiomia. |  |
Sirenia
| Name | Species | Member | Locality | Material | Notes | Image |
|---|---|---|---|---|---|---|
| Eosiren[59][60] | Eosiren imenti | Upper Sequence | Quarry O | Skull and ribs. | E. imenti is the most derived member of the genus Eosiren and had an elongated head. |  |
See also
References
- ^ a b c d e f g h i j k l m n o p q r s t u Rasmussen, D.T.; Olson, S.L.; Simons, E.L. (1987). "Fossil birds from the Oligocene Jebel Qatrani formation Fayum Province, Egypt". Smithsonian Contributions to Paleobiology. 62: 1–20.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am Kampouridis, P.; Hartung, J.; Augustin, F.J. (2023). "The Eocene–Oligocene Vertebrate Assemblages of the Fayum Depression, Egypt". Advances in Science, Technology & Innovation. doi:10.1007/978-3-030-95637-0_14.
- ^ a b c Seiffert Erik R. (Jan 2006). "Revised age estimates for the later Paleogene mammal faunas of Egypt and Oman". Proceedings of the National Academy of Sciences of the United States of America. 103 (13): 5000–5005. Bibcode:2006PNAS..103.5000S. doi:10.1073/pnas.0600689103. PMC 1458784. PMID 16549773.
- ^ a b c d e f g h i j Murray, A.M. (2004). "Late Eocene and early Oligocene teleost and associated ichthyofauna of the Jebel Qatrani Formation, Fayum, Egypt". Palaeontology. 47 (3): 711–724. doi:10.1111/j.0031-0239.2004.00384.x.
- ^ a b Rasmussen, D. T.; Simons, E.L.; Hertel, F.; Judd, A. (2001). "Hindlimb of a giant terrestrial bird from the upper Eocene, Fayum, Egypt". Palaeontology. 44 (2): 325–337. doi:10.1111/1475-4983.00182.
- ^ a b c d Gaffney, E.S.; Meylan, P.A.; Wood, R.C.; Simons, E.; de Almeida Campos, D. (2011). "Evolution of the side-necked turtles: the family Podocnemididae". Bulletin of the American Museum of Natural History. 350: 1–237. doi:10.1206/350.1. hdl:2246/6110. S2CID 83775718.
- ^ Pérez-García, A. (2017). "New information and establishment of a new genus for the Egyptian Paleogene turtle "Stereogenys" libyca (Podocnemididae, Erymnochelyinae)". Historical Biology: 1–10. doi:10.1080/08912963.2017.1374383.
- ^ a b Brochu, C.A.; Gingerich, P.D. (2000). "New tomistomine crocodylian from the middle Eocene (Bartonian) of Wadi Hitan, Fayum Province, Egypt" (PDF). CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN. 30 (10): 251–268.
- ^ Gaffney, E.S.; Deblieux, D.D.; Simons, E.L.; Sánchez-Villagra, M.R.; Meylan, P.A. (2002). "Redescription of the Skull of Dacquemys, a Podocnemidid Side-Necked Turtle from the Late Eocene of Egypt" (PDF). American Museum Novitates: 1–16. doi:10.1206/0003-0082(2002)372<0001:ROTSOD>2.0.CO;2.
- ^ Holroyd, P.A.; Parham, J.F. (2003). "The antiquity of African tortoises". Journal of Vertebrate Paleontology. 23 (3): 688–690. doi:10.1671/1870.
- ^ a b c d Seiffert, E.R. (2010). "The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt". Acta Palaeontologica Polonica. 55 (4): 599–616. doi:10.4202/app.2010.0023.
- ^ Simons, E.L.; Holroyd, P.A.; Bown, T.M. (1991). "Early tertiary elephant-shrews from Egypt and the origin of the Macroscelidea". Proceedings of the National Academy of Sciences. 88 (21): 9734–9737.
- ^ a b c Pickford, M. (2019). "Tiny Tenrecomorpha (Mammalia) from the Eocene of Black Crow, Namibia" (PDF). Communications of the Geological Survey of Namibia. 21: 15–25.
- ^ a b Sileem, A.H.; Sallam, H.M.; Hewaidy, A.G.A.; Gunnell, G.F.; Miller, E.R. (2015). "Anthracotheres (Mammalia, Artiodactyla) from the upper-most horizon of the Jebel Qatrani formation, latest Early Oligocene, Fayum depression, Egypt" (PDF). Egyptian Journal of Paleontology. 15: 1–11. ISSN 1687-4986.
- ^ a b c Holroyd, P.A.; Simons, E.L.; Bown, T.M.; Polly, P.D.; Kraus, M.J. (1996). "New records of terrestrial mammals from the upper Eocene Qasr el Sagha Formation, Fayum Depression, Egypt" (PDF). Palaeovertebrata. 25 (2–4): 175–192.
- ^ a b c Sileem, A.H.; Abu El‐Kheir, G.A. (2022). "Complete skull of Bothriogenys fraasi (Mammalia, Artiodactyla, Anthracotheriidae) from the Early Oligocene, Fayum, Egypt" (PDF). Geological Journal. doi:10.1002/gj.4574.
- ^ O'Leary, M.A.; Patel, B.A.; Coleman, M.N. (2012). "Endocranial petrosal anatomy of Bothriogenys (Mammalia, Artiodactyla, Anthracotheriidae), and petrosal volume and density comparisons among aquatic and terrestrial artiodactyls and outgroups" (PDF). Journal of Paleontology. 86 (1): 44–50. doi:10.1666/10-091.1.
- ^ Sileem, A.H.; Sallam, H.M.; Hewaidy, A.G.A.; Miller, E.R.; Gunnell, G.F. (2016). "A new anthracothere (Artiodactyla) from the early Oligocene, Fayum, Egypt, and the mystery of African 'Rhagatherium' solved". Journal of Paleontology. 90 (1): 170–181. doi:10.1017/jpa.2016.13.
- ^ Patricia A. Holroyd, Fabrice Lihoreau, Gregg F. Gunnell und Ellen R. Miller: Anthracotheriidae. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 843–851
- ^ a b c d e f Simmons, N.B.; Seiffert, E.R.; Gunnell, G.F. (2016). "A new family of large omnivorous bats (Mammalia, Chiroptera) from the Late Eocene of the Fayum Depression, Egypt, with comments on use of the name "Eochiroptera"" (PDF). American Museum Novitates (3857): 1–43. ISSN 0003-0082.
- ^ a b c Gunnell, G. F.; Simons, E. L.; Seiffert, E. R. (2008). "New bats (Mammalia: Chiroptera) from the late Eocene and early Oligocene, Fayum Depression, Egypt". Journal of Vertebrate Paleontology. 28 (1): 1–11. doi:10.1671/0272-4634(2008)28[1:nbmcft]2.0.co;2.
- ^ Gunnell, G.F.; Simmons, N.B.; Seiffert, E.R. (2014). "New Myzopodidae (Chiroptera) from the Late Paleogene of Egypt: Emended Family Diagnosis and Biogeographic Origins of Noctilionoidea". PLoS ONE. 9 (2). doi:10.1371/journal.pone.0086712.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Court, N. (1990). "Periotic anatomy of Arsinoitherium (Mammalia, Embrithopoda) and its phylogenetic implications". Journal of Vertebrate Paleontology. 10 (2): 170–182. doi:10.1080/02724634.1990.10011806.
- ^ Vialle, N.; Merzeraud, G.; Delmer, C.; Feist, M.; Jiquel, S.; Marivaux, L.; Ramdarshan, A.; Vianey-Liaud, M.; Essid, E.M.; Marzougui, W.; Ammar, H.K.; Tabuce, R. (2013). "Discovery of an Embrithopod Mammal (Arsinoitherium?) in the Late Eocene of Tunisia" (PDF). Journal of African Earth Sciences. 87: 86–92. doi:10.1016/j.jafrearsci.2013.07.010.
- ^ a b c d e f Borths, M.R.; Holroyd, P.A.; Seiffert, E.R. (2016). "Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia)". PeerJ. doi:10.7717/peerj.2639.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b c d e Osborn, H.F. (1909). "New carnivorous mammals from the Fayûm Oligocene, Egypt" (PDF). Bulletin of the American Museum of Natural History. 26: 415–424.
- ^ a b c d e f Holroyd, P.A. (1999). "New Pterodontinae (Creodonta: Hyaenodontidae) from the late Eocene-early Oligocene Jebel Qatrani Formation, Fayum province, Egypt". PaleoBios. 19 (2): 1–18.
- ^ Simons, E.L.; Gingerich, P.D. (1976). "A new species of Apterodon (Mammalia, Creodonta) from the upper Eocene Qasr el-Sagha Formation of Egypt". Postilla of the Peabody Museum of Natural History.
- ^ a b c Lange-Badré, B.; Böhme, M. (2005). "Apterodon intermedius, sp. nov., a new European creodont mammal from MP22 of Espenhain (Germany)". Annales de Paléontologie. 91 (4). doi:10.1016/j.annpal.2005.08.001.
- ^ a b c d e Borths, M.R.; Seiffert, E.R. (2017). "Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts". PLoS ONE. 12 (4). doi:10.1371/journal.pone.0173527.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b c d Borths, M.R.; Stevens, N.J. (2017). "The first hyaenodont from the late Oligocene Nsungwe Formation of Tanzania: Paleoecological insights into the Paleogene-Neogene carnivore transition". PLoS ONE. 12 (10). doi:10.1371/journal.pone.0185301.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b Morales, J.; Pickford, M. (2017). "New hyaenodonts (Ferae, Mammalia) from the early Miocene of Napak (Uganda), Koru (Kenya) and Grillental (Namibia)". Fossil Imprint. 73 (3–4): 332–359. doi:10.2478/if-2017-0019.
- ^ Solé, F.; Lhuillier, J.; Adaci, M.; Bensalah, M.; Mahboubi, M.; Tabuce, R. (2014). "The hyaenodontidans from the Gour Lazib area (? early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae" (PDF). Journal of Systematic Palaeontology. 12 (3): 303–322. doi:10.1080/14772019.2013.795196.
- ^ Andrews, C.W. (1906). "A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt". Order of the Trustees of the British Museum.
- ^ Morlo, M.; Bastl, K.; Wenhao, W.; Schaal, S.F.K. (2014). "The first species of Sinopa (Hyaenodontida, Mammalia) from outside of North America: implications for the history of the genus in the Eocene of Asia and North America". Palaeontology. 57 (1): 111–125. doi:10.1111/pala.12052.
- ^ a b c d e f g h i j k l m n Rasmussen, D.T.; Simons, E.L. (2000). "Ecomorphological diversity among Paleogene hyracoids (Mammalia): a new cursorial browser from the Fayum, Egypt" (PDF). Journal of Vertebrate Paleontology. 20 (1): 167–176. doi:10.1671/0272-4634(2000)020[0167:EDAPHM]2.0.CO;2.
- ^ De Blieux, D.D.; Simons, E.L. (2002). "Cranial and dental anatomy of Antilohyrax pectidens: a late Eocene hyracoid (Mammalia) from the Fayum, Egypt". Journal of Vertebrate Paleontology. 22 (1): 122–136. doi:10.1671/0272-4634(2002)022[0122:CADAOA]2.0.CO;2.
- ^ a b c d e f g h i j k l De Blieux, D.D.; Baumrind, M.R.; Simons, E.L.; Chatrath, P.S.; Meyer, G.E.; Attia, Y.S. (2006). "Sexual dimorphism of the internal mandibular chamber in Fayum Pliohyracidae (Mammalia)" (PDF). Journal of Vertebrate Paleontology. 26 (1): 160–169.
- ^ a b c d e Meyer, G.E. (1973). "A new Oligocene hyrax from the Jebel el Qatrani formation, Fayum, Egypt" (PDF). Peabody Museum of Natural History, Yale University.
- ^ a b c d e f g h i Tabuce, R. (2016). "A mandible of the hyracoid mammal Titanohyrax andrewsi in the collections of the Muséum National d'Histoire Naturelle, Paris (France) with a reassessment of the species". Palaeovertebrata. 40 (1). doi:10.18563/pv.40.1.e4.
- ^ a b c d e f g h i j Rasmussen, D. T.; Simons, E. L. (1988). "New Oligocene hyracoids from Egypt". Journal of Vertebrate Paleontology. 8 (1): 67–83. doi:10.1080/02724634.1988.10011684.
- ^ a b Matsumoto, H. (1922). "Megalohyrax, Andrews and Titanohyrax, gen. nov. A revision of the genera of hyracoids from the Fayum, Egypt". Proceedings of the Zoological Society (1921): 839–850.
- ^ a b c d Rasmussen, D.T.; Simons, E.L. (1991). "The Oldest Egyptian Hyracoids (Mammalia: Pliohyracidae): New Species of Saghatherium and Thyrohyrax from the Fayum". Neues Jahrbuch für Geologie und Paläontologie. 182 (2): 187–209. doi:10.1127/njgpa/182/1991/187.
- ^ Hooker, J.J.; Sánchez-Villagra, M.R.; Goin, F.J.; Simons, E.L.; Attia, Y.; Seiffert, E.R. (2008). "The origin of Afro‐Arabian 'didelphimorph'marsupials". Palaeontology. 51 (3): 635–648. doi:10.1111/j.1475-4983.2008.00779.x.
- ^ a b c d e f Al-Ashqar, SF; Seiffert, E.R.; de Vries, D.; El-Sayed, S.; Antar, M.S.; Sallam, H.M. (2021). "New phiocricetomyine rodents (Hystricognathi) from the Jebel Qatrani Formation, Fayum Depression, Egypt". PeerJ. 9. doi:10.7717/peerj.12074.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b c d e f g Sallam, H.M.; Seiffert, E.R.; Simons, E.L. (2012). "A basal phiomorph (Rodentia, Hystricognathi) from the late Eocene of the Fayum Depression, Egypt". Swiss Journal of Palaeontology. 131: 283–301. doi:10.1007/s13358-012-0039-6.
- ^ a b Sallam, H.M.; Seiffert, E.R. (2016). "New phiomorph rodents from the latest Eocene of Egypt, and the impact of Bayesian "clock"-based phylogenetic methods on estimates of basal hystricognath relationships and biochronology". PeerJ. 4. doi:10.7717/peerj.1717.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Sallam, H.M.; Seiffert, E.R.; Simons, E.L. (2011). "Craniodental Morphology and Systematics of a New Family of Hystricognathous Rodents (Gaudeamuridae) from the Late Eocene and Early Oligocene of Egypt". PLoS ONE. 6 (2). doi:10.1371/journal.pone.0016525.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Sallam, H.M.; Seiffert, E.R. (2020). "Revision of Oligocene 'Paraphiomys' and an origin for crown Thryonomyoidea (Rodentia: Hystricognathi: Phiomorpha) near the Oligocene–Miocene boundary in Africa" (PDF). Zoological Journal of the Linnean Society. 190 (1): 352–371.
- ^ Jaeger, J.J.; Marivaux, L.; Salem, M.; Bilal, A.A.; Benammi, M.; Chaimanee, Y.; Duringer, P.; Marandat, B.; Métais, E.; Schuster, M.; Valentin, X.; Brunet, M. (2010). "New rodent assemblages from the Eocene Dur At-Talah escarpment (Sahara of central Libya): systematic, biochronological, and palaeobiogeographical implications" (PDF). Zoological Journal of the Linnean Society. 160 (1): 195–213.
- ^ a b c Bown, T.M.; Simons, E.L. (1987). "New Oligocene Ptolemaiidae (Mammalia: ?Pantolesta) from the Jebel Qatrani Formation, Fayum Depression, Egypt". Journal of Vertebrate Paleontology. 7 (3): 311–324. doi:10.1080/02724634.1987.10011662.
- ^ a b c Simons, E.L.; Bown, T.M. (1995). "Ptolemaiida, a new order of Mammalia--with description of the cranium of Ptolemaia grangeri". Proc Natl Acad Sci U S A. 92 (8): 3269–73. doi:10.1073/pnas.92.8.3269. PMC 42147. PMID 11607526.
- ^ Osborn, H. F. (1908). "New fossil mammals from the Fayum Oligocene, Egypt". Bulletin of the American Museum of Natural History. 24: 265–272.
- ^ a b c d e f g h i Seiffert, E. R.; Simons, E. L. (2013). "Last of the oligopithecids? A dwarf species from the youngest primate-bearing level of the Jebel Qatrani Formation, northern Egypt". Journal of Human Evolution. 64 (3): 211–215. doi:10.1016/j.jhevol.2012.10.011.
- ^ a b c d e Simons, E. L.; Seiffert, E. R.; Chatrath, P. S.; Attia, Y. (2001). "Earliest Record of a Parapithecid Anthropoid from the Jebel Qatrani Formation, Northern Egypt". Folia Primatologica. 72 (6): 316–331. doi:10.1159/000052748.
- ^ a b Simons, E. L. (1992). "Diversity in the early Tertiary anthropoidean radiation in Africa". Proceedings of the National Academy of Sciences. 89 (22): 10743–10747. Bibcode:1992PNAS...8910743S. doi:10.1073/pnas.89.22.10743. PMC 50418. PMID 1438271.
- ^ a b Simons, E.L. (1989). "Description of two genera and species of late Eocene Anthropoidea from Egypt". Proceedings of the National Academy of Sciences. 86 (24): 9956–9960.
- ^ Simons, E.L.; Kay, R.F. (1983). "Qatrania, new basal anthropoid primate from the Fayum, Oligocene of Egypt". Nature. 304: 624–626.
- ^ Domning, D.P.; Gingerich, P.D.; Zalmout, I.S. (2010). "14: Sirenia". Cenozoic Mammals of Africa. pp. 147–160.
- ^ Domning, D.P.; Gingerich, P.D.; Simons, E.L.; Ankel-Simons, F.A. (1994). "A new early Oligocene dugongid (Mammalia, Sirenia) from Fayum Province, Egypt" (PDF). Contributions from the Museum of Paleontology The University of Michigan. 29 (4).