
Caudate nucleus
| Caudate nucleus | |
|---|---|
 Transverse Cut of Brain (Horizontal Section), basal ganglia is blue | |
| Details | |
| Identifiers | |
| Latin | nucleus caudatus |
| MeSH | D002421 |
| NeuroNames | 226 |
| NeuroLex ID | birnlex_1373 |
| TA98 | A14.1.09.502 |
| TA2 | 5561 |
| FMA | 61833 |
| Anatomical terms of neuroanatomy | |
The caudate nucleus is a nucleus located within the basal ganglia of the brains of many animal species. The caudate nucleus is an important part of the brain's learning and memory system.
Anatomy

The caudate nuclei are located near the center of the brain, sitting astride the thalamus. There is a caudate nucleus within each hemisphere of the brain. Individually, they resemble a C-shape structure with a wider "head" (caput in Latin) at the front, tapering to a "body" (corpus) and a "tail" (cauda). Sometimes a part of the caudate nucleus is referred to as the "knee" (genu).[1]

The head and body of the caudate nucleus form part of the floor of the anterior horn of the lateral ventricle. After the body travels briefly towards the back of the head, the tail curves back toward the anterior, forming the roof of the inferior horn of the lateral ventricle. This means that a coronal (on the same plane as the face) section that cuts through the tail will also cross the body and head of the caudate nucleus.
The caudate nucleus is related anatomically to a number of other structures. It is separated from the lenticular nucleus (made up of the globus pallidus and the putamen) by the anterior limb of the internal capsule. Together the caudate and putamen form the dorsal striatum.
Neurochemistry
The caudate nucleus is highly innervated by dopamine neurons. These neurons originate mainly from the ventral tegmental area (VTA) and the substantia nigra pars compacta (SNc). There are also additional inputs from various association cortices.
Physiology
Learning and memory
Historically, the basal ganglia as a whole have been implicated in higher-order motor control.[2] The caudate nucleus was initially thought to primarily be involved with control of voluntary movement. More recently, it has been demonstrated that the caudate is highly involved in learning and memory,[3] particularly regarding feedback processing.[4] In general, it has been demonstrated that neural activity will be present within the caudate while an individual is receiving feedback. People with hyperthymesia appear to have slight increases in the sizes of the caudate nucleus as well as of the temporal lobe of the cortex.[5]
Emotion
The caudate nucleus has been implicated in responses to visual beauty, and has been suggested as one of the "neural correlates of romantic love".[6][7]
Language comprehension
The left caudate in particular has been suggested to have a relationship with the thalamus that governs the comprehension and articulation of words as they are switched between languages.[8][9]
Threshold control
The brain contains large collections of neurons reciprocally connected by excitatory synapses, thus forming large network of elements with positive feedback. It is difficult to see how such a system can operate without some mechanism to prevent explosive activation. There is some indirect evidence[10] that the caudate may perform this regulatory role by measuring the general activity of cerebral cortex and controlling the threshold potential.
Role in obsessive compulsive disorder
It has been theorized that the caudate nucleus may be dysfunctional in persons with obsessive compulsive disorder (OCD), in that it may perhaps be unable to properly regulate the transmission of information regarding worrying events or ideas between the thalamus and the orbitofrontal cortex.
A neuroimaging study with positron emission tomography found that the right caudate nucleus had the largest change in glucose metabolism after patients had been treated with paroxetine.[11] Recent SDM meta-analyses of voxel-based morphometry studies comparing people with OCD and healthy controls have found people with OCD to have increased grey matter volumes in bilateral lenticular nuclei, extending to the caudate nuclei, while decreased grey matter volumes in bilateral dorsal medial frontal/anterior cingulate gyri.[12][13] These findings contrast with those in people with other anxiety disorders, who evince decreased (rather than increased) grey matter volumes in bilateral lenticular / caudate nuclei, while also decreased grey matter volumes in bilateral dorsal medial frontal/anterior cingulate gyri.[13]
Additional images
-
 Diagram of hippocampus
Diagram of hippocampus -
 Velum interpositum.
Velum interpositum. -
 Schematic representation of the chief ganglionic categories (I to V).
Schematic representation of the chief ganglionic categories (I to V). -
 Superficial dissection of brain-stem. Lateral view.
Superficial dissection of brain-stem. Lateral view. -
 Dissection of brain-stem. Lateral view.
Dissection of brain-stem. Lateral view. -
 Deep dissection of brain-stem. Lateral view.
Deep dissection of brain-stem. Lateral view. -
 Deep dissection of brain-stem. Lateral view.
Deep dissection of brain-stem. Lateral view. -
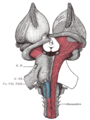 Superficial dissection of brain-stem. Ventral view.
Superficial dissection of brain-stem. Ventral view. -
 Dissection of brain-stem. Dorsal view.
Dissection of brain-stem. Dorsal view. -
 Dissection showing the ventricles of the brain.
Dissection showing the ventricles of the brain. -
 Coronal section of brain immediately in front of pons.
Coronal section of brain immediately in front of pons. -
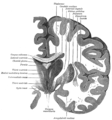 Coronal section of brain through intermediate mass of third ventricle.
Coronal section of brain through intermediate mass of third ventricle. -
 Coronal section of lateral and third ventricles.
Coronal section of lateral and third ventricles. -
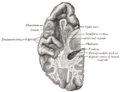 Section of brain showing upper surface of temporal lobe.
Section of brain showing upper surface of temporal lobe. -
 Central part and anterior and posterior cornua of lateral ventricles exposed from above.
Central part and anterior and posterior cornua of lateral ventricles exposed from above. -

-
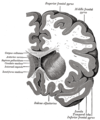 Coronal section through anterior cornua of lateral ventricles.
Coronal section through anterior cornua of lateral ventricles. -
 Coronal section of brain through anterior commissure.
Coronal section of brain through anterior commissure. -
 Coronal section of inferior horn of lateral ventricle.
Coronal section of inferior horn of lateral ventricle. -
 Human brain left dissected midsagittal view
Human brain left dissected midsagittal view -
 Human brain right dissected lateral view
Human brain right dissected lateral view -
 purple=caudate nucleus and putamen, orange=thalamus
purple=caudate nucleus and putamen, orange=thalamus -
 Caudate nucleus
Caudate nucleus -
 Caudate nucleus
Caudate nucleus
.jpg)























References
- ^ E. H. Yeterian, D. N. Pandya, "Corticostriatal connections of extrastriate visual areas in rhesus monkeys," The Journal of Comparative Neurology 352(3):436-457, 1995. PMID: 7706560
- ^ S. A. Kinnier Wilson (1914). "An experimental research into the anatomy of the corpus striatum". Brain. 36 (3–4): 427–492. doi:10.1093/brain/36.3-4.427.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ Graybiel AM (2005) The basal ganglia: learning new tricks and loving it. Curr Opin Neurobiol 15:638-644.
- ^ Packard MG, Knowlton BJ (2002) Learning and memory functions of the Basal Ganglia. Annu Rev Neurosci 25:563-593.
- ^ "The Gift of Endless Memory". CBS News. 16 December 2010.
- ^ Tomohiro Ishizu and Semir Zeki (2011). Warrant, Eric James (ed.). "Toward A Brain-Based Theory of Beauty". PLoS ONE. 6 (7): 1–10. doi:10.1371/journal.pone.0021852.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: unflagged free DOI (link) - ^ Arthur Aron, Helen Fisher, Debra J. Mashek, Greg Strong, Haifang Li, and Lucy L. Brown (2005). "Reward, Motivation, and Emotion Systems Associated With Early-Stage Intense Romantic Love". J. Neurophysiol. 94: 327–337. doi:10.1152/jn.00838.2004.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ "How bilingual brains switch between tongues" at newscientist.com
- ^ "Language Control in the Bilingual Brain " at sciencemag.org
- ^ Braitenberg V. (1984)Vehicles. Experiments in synthetic psychology.
- ^ Elsebet S. Hansen, Steen Hasselbalch, Ian Law; Tom G. Bolwig (2002). "The caudate nucleus in obsessive-compulsive disorder. Reduced metabolism following treatment with paroxetine: a PET study". International Journal of Neuropsychopharmacology. 5 (1): 1–10. doi:10.1017/S1461145701002681. PMID 12057027.
{{cite journal}}: Check date values in:|year=(help)CS1 maint: multiple names: authors list (link) - ^ Radua, Joaquim; Mataix-Cols, David (2009). "Voxel-wise meta-analysis of grey matter changes in obsessive–compulsive disorder". British Journal of Psychiatry. 195 (5): 393–402. doi:10.1192/bjp.bp.108.055046. PMID 19880927.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ a b Radua, Joaquim; van den Heuvel, Odile A.; Surguladze, Simon; Mataix-Cols, David (5 July 2010). "Meta-analytical comparison of voxel-based morphometry studies in obsessive-compulsive disorder vs other anxiety disorders". Archives of General Psychiatry. 67 (7): 701–711. doi:10.1001/archgenpsychiatry.2010.70. PMID 20603451.
{kind=link}