
Organ of Corti
This article needs additional citations for verification. (December 2014) |
| Organ of Corti | |
|---|---|
 A cross section of the cochlea illustrating the organ of Corti | |
| Details | |
| Part of | Cochlea of the inner ear |
| Identifiers | |
| Latin | organum spirale |
| MeSH | D009925 |
| NeuroLex ID | birnlex_2526 |
| TA98 | A15.3.03.121 |
| TA2 | 7035 |
| FMA | 75715 |
| Anatomical terminology | |
The organ of Corti, or spiral organ, is the receptor organ for hearing and is located in the mammalian cochlea. This highly varied strip of epithelial cells allows for transduction of auditory signals into nerve impulses' action potential.[1] Transduction occurs through vibrations of structures in the inner ear causing displacement of cochlear fluid and movement of hair cells at the organ of Corti to produce electrochemical signals.[2]
Italian anatomist Alfonso Giacomo Gaspare Corti (1822–1876) discovered the organ of Corti in 1851.[3] The structure evolved from the basilar papilla and is crucial for mechanotransduction in mammals.
Structure
[edit]
The organ of Corti is located in the scala media of the cochlea of the inner ear between the vestibular duct and the tympanic duct and is composed of mechanosensory cells, known as hair cells.[2] Strategically positioned on the basilar membrane of the organ of Corti are three rows of outer hair cells (OHCs) and one row of inner hair cells (IHCs).[4] Surrounding these hair cells are supporting cells: Deiters cells, also called phalangeal cells, which have a close relation with the OHCs, and pillar cells, which separate and support both the OHCs and the IHCs.[4]
Projecting from the tops of the hair cells are tiny finger-like projections called stereocilia, which are arranged in a graduated fashion with the shortest stereocilia on the outer rows and the longest in the center. This gradation is thought to be the most important anatomic feature of the organ of Corti because this allows the sensory cells superior tuning capability.[5]
If the cochlea were uncoiled, it would roll out to be about 33 mm long in women and 34 mm in men, with about 2.28 mm of standard deviation for the population.[6] The cochlea is also tonotopically organized, meaning that different frequencies of sound waves interact with different locations on the structure. The base of the cochlea, closest to the outer ear, is the most stiff and narrow and is where the high-frequency sounds are transduced. The apex, or top, of the cochlea is wider and much more flexible and loose and functions as the transduction site for low-frequency sounds.[7]
Function
[edit]
The function of the organ of Corti is to convert (transduce) sounds into electrical signals that can be transmitted to the brainstem through the auditory nerve.[2] It is the auricle and middle ear that act as mechanical transformers and amplifiers so that the sound waves end up with amplitudes 22 times greater than when they entered the ear.
Auditory transduction
[edit]In normal hearing, the majority of the auditory signals that reach the organ of Corti in the first place come from the outer ear. Sound waves enter through the auditory canal and vibrate the tympanic membrane, also known as the eardrum, which vibrates three small bones called the ossicles. As a result, the attached oval window moves and causes movement of the round window, which leads to displacement of the cochlear fluid.[8] However, the stimulation can happen also via direct vibration of the cochlea from the skull. The latter is referred to as Bone Conduction (or BC) hearing, as complementary to the first one described, which is instead called Air Conduction (or AC) hearing. Both AC and BC stimulate the basilar membrane in the same way (Békésy, G.v., Experiments in Hearing. 1960).
The basilar membrane on the tympanic duct presses against the hair cells of the organ as perilymphatic pressure waves pass. The stereocilia atop the IHCs move with this fluid displacement and in response their cation, or positive ion selective, channels are pulled open by cadherin structures called tip links that connect adjacent stereocilia.[9] The organ of Corti, surrounded in potassium-rich fluid endolymph, lies on the basilar membrane at the base of the scala media. Under the organ of Corti is the scala tympani and above it, the scala vestibuli. Both structures exist in a low potassium fluid called perilymph.[8] Because those stereocilia are in the midst of a high concentration of potassium, once their cation channels are pulled open, potassium ions as well as calcium ions flow into the top of the hair cell. With this influx of positive ions the IHC becomes depolarized, opening voltage-gated calcium channels at the basolateral region of the hair cells and triggering the release of the neurotransmitter glutamate. An electrical signal is then sent through the auditory nerve and into the auditory cortex of the brain as a neural message.
Cochlear amplification
[edit]The organ of Corti is also capable of modulating the auditory signal.[7] The outer hair cells (OHCs) can amplify the signal through a process called electromotility where they increase movement of the basilar and tectorial membranes and therefore increase deflection of stereocilia in the IHCs.[8][10][11]
A crucial piece to this cochlear amplification is the motor protein prestin, which changes shape based on the voltage potential inside of the hair cell. When the cell is depolarized, prestin shortens, and because it is located on the membrane of OHCs it then pulls on the basilar membrane and increasing how much the membrane is deflected, creating a more intense effect on the inner hair cells (IHCs). When the cell hyperpolarizes prestin lengthens and eases tension on the IHCs, which decreases the neural impulses to the brain. In this way, the hair cell itself is able to modify the auditory signal before it even reaches the brain.
Development
[edit]The organ of Corti, in between the scala tympani and the scala media, develops after the formation and growth of the cochlear duct.[7] The inner and outer hair cells then differentiate into their appropriate positions and are followed by the organization of the supporting cells. The topology of the supporting cells lends itself to the actual mechanical properties that are needed for the highly specialized sound-induced movements within the organ of Corti.[7]
Development and growth of the organ of Corti relies on specific genes, many of which have been identified in previous research (SOX2, GATA3, EYA1, FOXG1, BMP4, RAC1, and more),[7] to undergo such differentiation. Specifically, the cochlear duct growth and the formation of hair cells within the organ of Corti.
Mutations in the genes expressed in or near the organ of Corti before the differentiation of hair cells will result in a disruption in the differentiation, and potential malfunction of, the organ of Corti.
Clinical significance
[edit]Hearing loss
[edit]The organ of Corti can be damaged by excessive sound levels, leading to noise-induced impairment.[12]
The most common kind of hearing impairment, sensorineural hearing loss, includes as one major cause the reduction of function in the organ of Corti. Specifically, the active amplification function of the outer hair cells is very sensitive to damage from exposure to trauma from overly-loud sounds or to certain ototoxic drugs. Once outer hair cells are damaged, they do not regenerate, and the result is a loss of sensitivity and an abnormally large growth of loudness (known as recruitment) in the part of the spectrum that the damaged cells serve.[13]
While hearing loss has always been considered irreversible in mammals, fish and birds routinely repair such damage. A 2013 study has shown that the use of particular drugs may reactivate genes normally expressed only during hair cell development. The research was carried out at Harvard Medical School, Massachusetts Eye and Ear, and the Keio University School of Medicine in Japan.[14][15]
Additional images
[edit]-
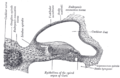 Transverse section of the cochlear duct of a fetal cat.
Transverse section of the cochlear duct of a fetal cat. -
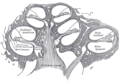 Diagrammatic longitudinal section of the cochlea
Diagrammatic longitudinal section of the cochlea -
 Floor of ductus cochlearis
Floor of ductus cochlearis -
 Limbus laminæ spiralis and membrana basilaris
Limbus laminæ spiralis and membrana basilaris -
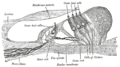 Section through the spiral organ of Corti (magnified)
Section through the spiral organ of Corti (magnified)




Notes
[edit]- ^ Hudspeth, A (2014). "Integrating the active process of hair cells with cochlear function". Nature Reviews Neuroscience. 15 (9): 600–614. doi:10.1038/nrn3786. PMID 25096182. S2CID 3716179.
- ^ a b c The Ear Pujol, R., Irving, S., 2013
- ^ Betlejewski, S (2008). "Science and life – the history of Marquis Alfonso Corti". Otolaryngologia Polska. 62 (3): 344–347. doi:10.1016/S0030-6657(08)70268-3. PMID 18652163.
- ^ a b Malgrange, B; Van de Water, T.R; Nguyen, L; Moonen, G; Lefebvre, P.P (2002). "Epithelial supporting cells can differentiate into outer hair cells and Deiters' cells in the cultured organ of Corti". Cellular and Molecular Life Sciences. 59 (10): 1744–1757. doi:10.1007/pl00012502. PMC 11337542. PMID 12475185. S2CID 2962483.
- ^ Lim, D (1986). "Functional structure of the organ of Corti: a review". Hearing Research. 22 (1–3): 117–146. doi:10.1016/0378-5955(86)90089-4. PMID 3525482. S2CID 4764624.
- ^ Miller, J. D. (2007). "Sex differences in the length of the organ of Corti in humans". The Journal of the Acoustical Society of America. 121 (4): EL151-5. Bibcode:2007ASAJ..121L.151M. doi:10.1121/1.2710746. PMID 17471760.
- ^ a b c d e Fritzsch, B; Jahan, I; Pan, N; Kers, J; Duncan, J; Kopecky, B (2012). "Dissecting the molecular basis of organ of Corti development: where are we now?". Hearing Research. 276 (1–2): 16–26. doi:10.1016/j.heares.2011.01.007. PMC 3097286. PMID 21256948.
- ^ a b c Nichols, J.G; Martin, A.R.; Fuchs, P.A; Brown, D.A; Diamond, M.E; Weisblat, D.A (2012). From Neuron to Brain, 5th Edition. Sunderland, MA: Sinauer Associates, Inc. pp. 456–459. ISBN 978-0-87893-609-0.
- ^ Müller, Ulrich; Gillespie, Peter G.; Williams, David S.; Reynolds, Anna; Dumont, Rachel A.; Lillo, Concepcion; Siemens, Jan (April 2004). "Cadherin 23 is a component of the tip link in hair-cell stereocilia". Nature. 428 (6986): 950–955. Bibcode:2004Natur.428..950S. doi:10.1038/nature02483. ISSN 1476-4687. PMID 15057245. S2CID 3506274.
- ^ Ashmore, Jonathan Felix (1987). "A fast motile response in guinea-pig outer hair cells: the cellular basis of the cochlear amplifier". The Journal of Physiology. 388 (1): 323–347. doi:10.1113/jphysiol.1987.sp016617. ISSN 1469-7793. PMC 1192551. PMID 3656195.

- ^ Ashmore, Jonathan (2008). "Cochlear Outer Hair Cell Motility". Physiological Reviews. 88 (1): 173–210. doi:10.1152/physrev.00044.2006. ISSN 0031-9333. PMID 18195086. S2CID 17722638.
- ^ Lim, David J. (March 1986). "Effects of noise and ototoxic drugs at the cellular level in the cochlea: A review". American Journal of Otolaryngology. 7 (2): 73–99. doi:10.1016/S0196-0709(86)80037-0. PMID 3515985.
- ^ Robert A. Dobie (2001). Medical-Legal Evaluation of Hearing Loss. Thomson Delmar Learning. ISBN 0-7693-0052-9.
- ^ "Cochlear hair cells - Beyond the Dish". wordpress.com.
- ^ Askew, Charles; Rochat, Cylia; Pan, Bifeng; Asai, Yukako; Ahmed, Hena; Child, Erin; Schneider, Bernard L.; Aebischer, Patrick; Holt, Jeffrey R. (8 July 2015). "Tmc gene therapy restores auditory function in deaf mice". Science Translational Medicine. 7 (295): 295ra108. doi:10.1126/scitranslmed.aab1996. ISSN 1946-6234. PMC 7298700. PMID 26157030.
References
[edit]- Corti A (1851). "Recherches sur l'organe de l'ouïe des mammiferes". Zeitschrift für Wissenschaftliche Zoologie. 3: 106–169.
- Fritzsch B, Jahan I, Pan N, Kersigo J, Duncan J, Kopecky B (2012). "Dissecting the molecular basis of organ of Corti development: where are we now?". Hearing Research. 276 (1–2): 16–26. doi:10.1016/j.heares.2011.01.007. PMC 3097286. PMID 21256948.
History. (n.d.).
- Hudspeth A (2014). "Integrating the active process of hair cells with cochlear function". Nature Reviews Neuroscience. 15 (9): 600–614. doi:10.1038/nrn3786. PMID 25096182. S2CID 3716179.
- Lim D (1986). "Functional structure of the organ of Corti: a review". Hearing Research. 22 (1–3): 117–146. doi:10.1016/0378-5955(86)90089-4. PMID 3525482. S2CID 4764624.
- Malgrange B, Thiry M, Van de Water TR, Nguyen L, Moonen G, Lefebvre PP (2002). "Epithelial supporting cells can differentiate into outer hair cells and Deiters' cells in the cultured organ of Corti". Cellular and Molecular Life Sciences. 59 (10): 1744–1757. doi:10.1007/PL00012502. PMC 11337542. PMID 12475185. S2CID 2962483.
- Nicholls, J. G., Martin, A. R., Fuchs, P. A., Brown, D. A., Diamond, M. E., & Weisblat, D. A. (2012). From Neuron to Brain (5th ed., pp. 456–459). Sunderland, MA: Sinauer Associates, Inc.
- Pritchard U. "On the organ of Corti in mammals". 2 March 1876, Proceedings of the Royal Society of London, volume 24, pp. 346–52 OCLC 1778190
- Pujol, R., & Irving, S. (2013). The Ear.
External links
[edit]- Dissecting the molecular basis of organ of Corti development PMC 3097286
- Organ of Corti 3D animation
- http://lobe.ibme.utoronto.ca/presentations/OHC_Electromotility/sld005.htm Archived 2005-03-30 at the Wayback Machine Diagram at University of Toronto
- http://mayoresearch.mayo.edu/mayo/research/ent_research/images/image02.gif Archived 2007-03-05 at the Wayback Machine Diagram at Mayo
- http://www.iurc.montp.inserm.fr/cric51/audition/english/corti/fcorti.htm Archived 2006-11-30 at the Wayback Machine at University of Montpellier 1
{kind=link}
{kind=link}