:The suggested change would make the diagram consistent with every neuroscience text I've ever used. [[User:Saccade|kraemer]] 00:17, 5 October 2007 (UTC)
:The suggested change would make the diagram consistent with every neuroscience text I've ever used. [[User:Saccade|kraemer]] 00:17, 5 October 2007 (UTC)
: Yes, you are both right. This is my diagram, and it is, in fact incorrect. The batteries should, in fact, be in series with the resistors. I've been meaning to fix it for a while, and just did not get around to it. I'll so it soon. [[User:Synaptidude|Synaptidude]]
The traditional current circulation is from the + pole to negative one. The real current circulation is from negative pole to positive. The T4 phase of the picture shows a green arrow (circulation) that doesn't respect the rules used in the previous phases.--[[User:Somasimple|Somasimple]] 06:50, 9 October 2007 (UTC)
The traditional current circulation is from the + pole to negative one. The real current circulation is from negative pole to positive. The T4 phase of the picture shows a green arrow (circulation) that doesn't respect the rules used in the previous phases.--[[User:Somasimple|Somasimple]] 06:50, 9 October 2007 (UTC)
This article is within the scope of WikiProject Spoken Wikipedia, a collaborative effort to improve the coverage of articles that are spoken on Wikipedia. If you would like to participate, please visit the project page, where you can join the discussion and see a list of open tasks.Spoken WikipediaWikipedia:WikiProject Spoken WikipediaTemplate:WikiProject Spoken WikipediaSpoken Wikipedia articles
"An action potential is an electrical pulse that changes the voltage (potential) across a cell membrane. In
muscle and nerve cells, this rapid change in voltage leads to an action such as muscular contraction or
neurotransmitter release. These actions are usually a consequence of calcium ions entering the cell
during the rapid change in voltage. "
I think any definition has to include the idea of propagation, if not the idea of a wave, which is the mode of propagation in the first kind of action potential most people think of, which is in nerves. It's just wrong that calcium "usually" causes an AP. According to the Hodgkin-Huxley theory, based on the way things work in the giant axon of the squid, all you need are sodium channels and potassium channels. Calcium is important in coupling excitation (the AP) to contraction in muscle, but at the neuromuscular junction its acetylcholine receptors that initiate the action potential, and these will predominantly be carrying sodium into the cell. I suspect they don't permit calcium at all.
I don't think the text as it is quoted is actually incorrect, but it sure could be worded better. The way I read that, the calcium influx is referring to that which cases muscular contraction or transmitter release, not the action potential itself. But the wording makes it ambiguous. Sodium and potassium are, indeed, the major players in most action potentials (cardiac action potentials, eg., have a significant calcium component). And just for point of trivia, the usual voltage-gated sodium channel is not 100% selective for sodium ions. Around 5% of the conductance is carried by calcium ions. Synaptidude23:43, 18 November 2005 (UTC)[reply]
Maybe I'm being pedantic here, but if you look at the phrase "action potential", it is clearly a noun. It should refer to the level of potential necessary to incite action (or some similar aspect of the amount of potential), not the process of changing potential. Is all of neurochemistry suffering a semantic deficit, or just this article? p.s. I'm coming from an electronics background, where potential clearly means "voltage" -- which, in this case, it literally does, so I think I'm standing on solid semantic ground. 12.210.113.23514:20, 1 June 2006 (UTC)[reply]
Picture
What does the cartoon of the phospholipid membrane have to do with an action potential? There aren't even any ion channels in the drawing, as far as I could tell. Could someone find a nice graph of an action potential (i.e. a voltage-time graph showing membrane potential increasing to threshold, then firing, then a refractory period)?Sayeth 19:52, Jul 29, 2004 (UTC)
I agree the top picture should be a voltage-time graph. Perhaps when one is found the membrane image could move further down the article? Richard Taylor
Thanks for the new caption, Richard. It makes the drawing much more relevant, but I agree that there's still some improvement possible with a second picture.Sayeth 19:52, Jul 29, 2004 (UTC)
The membrane was uploaded by me in search of a good image for the article, since it was listed as missing an image on Wikipedia talk:Featured article candidates. Apologies if it is only loosely related, and thanks for improving the caption, Richard. Following a request on Wikipedia:Requested pictures, I have created another image from scratch with the voltage, based on images found online at other locations. Hopefully this is more what you had in mind. (I am not very familiar with the topic) Happy editing -- Chris 73 | Talk 08:19, 30 Jul 2004 (UTC)
I had a few minutes of spare time and decided to redraw Chris' image in Adobe Illustrator. It's basically the same image with some antialiasing and smaller font sizes for readability. Would anyone object if I replaced the current image with my version? --Diberri | Talk 17:22, Jul 30, 2004 (UTC)
Nevermind my asking for permission. I decided to be bold and make the change. --Diberri | Talk 17:49, Jul 30, 2004 (UTC)
No problems. Looks good, too. -- Chris 73 | Talk 00:29, 31 Jul 2004 (UTC)
Is the refractory period a) that long and b) is the hyperpolarisation so "hyper" (ie does it go so negative?). I thougth that both were smaller. Batmanand 14:16, 22 Jun 2005 (UTC)
I agree the hyperpolarisation should not be that hyper. Somewhere between 10 and 15 mv would be more accurate. As far as the duration is concerned i think it should be longer, at least 4 ms. David D. 15:13, 22 Jun 2005 (UTC)
I'll add that for the refractory period any time from 1 -15 ms is technically correct since it does vary depending on physiological or developmental context. David D. 16:46, 23 Jun 2005 (UTC)
Lack of references
Hi this article no longer meets the criteria for a featured article because it does not cite its sources. Please help fix this so that all featured articles can meet the same standards. Best would be the most trusted resources in the field being added, some print resources especially, but also online references are better than none. Those sources would likely help with good material to further improve the article anyway. - Taxman 23:00, Oct 26, 2004 (UTC)
I added two of the most common neuroscience textbooks (Kandel and Bear) as references. If anyone who contributed to this article had more specific references in mind, please add them. Sayeth 17:42, Nov 1, 2004 (UTC)
Would you say you are very confident that the material in those books agrees with what is here? Otherwise that is potentially dishonest to list them as references. - Taxman 19:12, Nov 1, 2004 (UTC)
I'm familiar with the contents of Kandel, X and Schwartz. Yes, the content of that one aggrees with what is said here. I've not read Bear (although he did once hit on my girlfriend ;-), but this is all such "textbook stuff" that I'm sure Bear also supports what is in here.
I've also added references to the primary literature: the original HH papers and also an updated look at HH-models. Synaptidude 5 July 2005 20:58 (UTC)
Correct statements?
The first paragraph reads:
"They [AP's] set the pace of thought and action, constrain the sizes of evolving anatomies and enable centralized control and coordination of organs and tissues. Non-propagating action potentials occur also in some plants."
What is the evidence that the speed of an action potential contrains the size of an organism or organ? It is not discussed in the article.
Also, there are propagating action potentials in plants. They can move along the phloem cell membranes similar to nerves. A good example of this is the sensitive mimosa plant. It's leaflets close in response to touch and the movement can be seen to propagate down the stem. This is due to motor cells sequentially releasing water due to the change in membrane potential as the action potential progress' down the stem. This was observed and published by Charles Darwin in his less famous book The power of movement in plants. John Murray, London, 1880.
There are two big differences between plant and animal action potentials: 1) plant AP's are much slower; 2) plant AP's are generated with K+ and anions, unlike animals that use K+ and Na+ ions. David D.15:32, 3 May 2005 (UTC)[reply]
"constrain the sizes of evolving anatomies" Maybe there could be a section in the article that cites some references for research dealing with evolutionary constraints that involve action potential generation and propagation. Some textbooks contain statements about the large amount of energy required to support neuronal electrical activity. Some neurobiology texts discuss constraints on brain shape and size that are related to the speed of action potential propagation and how axon diameter and myelination contribute to that velocity. Some people have speculated about the possibility that large animals might be constrained in their body size by the speed of action potential propagation. Memenen02:12, 6 May 2005 (UTC)[reply]
You know, you could argue for days on end about chicken/egg aspects of evolutionary changes. I'm completely unfamiliar with any literature on action potentials and evolution, and yet it would seem obvious that the relationship between the two will be complex, probably showing causality in both directions (i.e. action potential propagation limits body size at the same time that body size limits axon diameter. Since evolutionary statements such as the one made in this article are often just informed speculation anyway, this whole problem can be (and has been) fixed by using the all-purpose scientist solution: "when you've got a feasible idea that you can't prove, say "may". Synaptidude 5 July 2005 21:05 (UTC)
New labels on voltage-time graph of AP
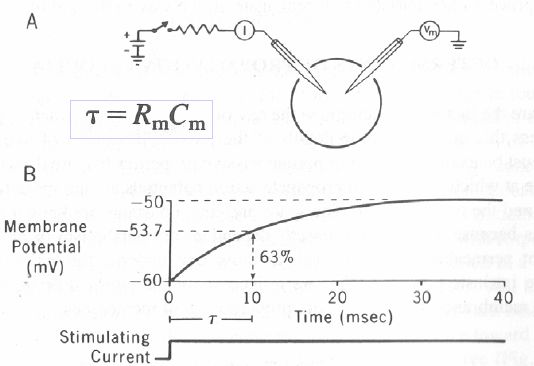
I relabeled the graph showing the voltage-time relationships of the action potential. The former labels were slightly misleading in the following ways. 1) "Depolarization" and "Hyperpolarization" have been replaced with "Rising" and "Falling Phase". First, these are the correct terminologies in use by electrophysiologists. Second, the Rising Phase, once it crosses 0 mV is actually hyperpolarizing (see hyperpolarization. Likewise, the falling phase before it reaches 0 mV is actually depolarizing (see depolarization). This is why electrophysiologists don't use those terms to describe the rising and falling phases (each is a mixed hyper/depolarization). 2) The label "Refractory Period" has been replaced by "Undershoot". This is because the label incorrectly implied that the undershoot of membrane potental at the end of the action potential is the basis for the refractory period. The basis for the refractory period is actually a change in the action potential threshold caused by sodium channel inactivation (which has nothing directly to do with the undershoot). The undershoot is actually too large in this diagram, but I decided not to fix this at this time. According to the voltage scale used, the undershoot would go to approx. -100 mV. This is below Ek, and therefore physically impossible. 3)I've added ~ marks to the voltage scale of the y-axis. This is to indicate that the values of resting, threshold and peak potentials are approximate. For example, neurons rest anywhere from -80 to -55 mV. Thresholds and peaks vary similarly. 4) I've added a label for "overshoot", that is, the part of the action potential that overshoots 0 mV.
Synaptidude 5 July 2005 18:59 (UTC)
Looks good. We discussed the magnitude of the hyperpolarisation above too. It is clearly way too much in that figure. David D. 5 July 2005 19:16 (UTC)
Yes, thanks. I see that now. I guess it's time to do something about it. Synaptidude 5 July 2005 21:10 (UTC)
Progagation and saltutory conduction
I'll have to check, but I think it was more likely Cole than H&H who "discovered" the propagating nature of action potentials. I think one could actually make a pretty strong argument for Sherington, who certainly knew that nerve impulses were propagated, although he didn't know specifically about action potentials per se
First Figure Parts A and B
I suggest putting Part A and Part B of the first figure like this:
A
B
As it is now, it is leaving too little room for the main text along the left side of the page. --Memenen 9 July 2005 16:52 (UTC)
Yes, the image is far to wide. The article looks like crap.
A more "complex" action potential.Some action potential are complex, with extra "bumps", "humps" and distinctive features in the voltage vs time graph. These extra features are usually due to specialized ion channels (such as calcium channels) and ion channel regulation processes that are important for certain physiological functions. An example is the cardiac action potential. --Memenen17:30, 18 August 2005 (UTC)[reply]
Reading the suggestions in the featured review section, I have undertaken some changes (hope they are improvments)
1) I've re-written the introduction to make it more general and accurate. The accuracy part is the the "sets the pace of thought" statement. I have removed this because there is no evidence for this. Most current thought is that the pace of thought is set by the speed of neural osicillators that are made of networks of many neurons. While action potentials certainly subserve parts of the function of these oscillators, they are not the rate-limiting steps. It would be no more accurate to say that the frequencies coming from a radio speaker are set by the speed of electron travel through copper. Synaptidude00:11, 22 November 2005 (UTC)[reply]
2) made some changes to clarify and more accurately reflect how the action potential is initiated
3) Added a section of "Why" the body uses action potentials
I've pared down a lot of the information that's been added over the last few months. There were quite a few asides not relevant to action potentials, per se, so I've either deleted them or (as was the case with the discussion of the squid giant axon) moved them to the relevant article. --David Iberri (talk) 19:53, 22 November 2005 (UTC)[reply]
David - most of what you added, or rather, subtracted, looks good to me, although much of your paring was of things I added only yesterday. But for the most part, I think you kept the gist of what I was trying to add, while at the same time curring down on my wordiness - which is just fine with me. The only part (so far) where I might disagree is in the "why action potential" portion I added yesterday. I think it's important to put something in the article about the advantages conferred by the action potential mechanism to cellular commication - but perhaps more skillfully than my attempt. Synaptidude23:01, 22 November 2005 (UTC)[reply]
My apologies for removing the "why an action potential?" section. I'd just copyedited the section on saltatory propagation and I misinterpreted your "why" text as describing the evolutionary advangates of myelin. As this clearly isn't the case and since the article could really use a section discussing the advantages conferred by action potentials, I've re-inserted the section. I fancy renaming it to eliminate the question mark, though I can't seem to do any better. --David Iberri (talk) 14:30, 23 November 2005 (UTC)[reply]
I flipped over to this Talk page to make some witty comment about how in depth this article is, and then I noticed it's a featured article...someone really knows their stuff heh
This review resulted in the passing of a new version of the article. Some minor extant issues remain unsolved, including the large diagram in the lead and the second paragraph of "control of timing".
This was the version promoted in February of 2004, and these are the changes made since. Article has been completely rewritten. The new version is quite a bit longer, and appears to be more comprehensive. The only significant issue I see is the diagram next to the lead, which makes for some very bad formatting (the lead is squeezed into really, really short lines), and the diagram seems a bit complex for the lead anyway (though I recognize this was likely done because no real image could be used for this article). Also, I don't like the section heading "Noteworthy characteristics of the action potential", though the content there seems fine. Absolutely no external links is not ideal, though there may be nothing worth linking to on such an esoteric topic. I'm also unsure about the "Related topics" section. All in all, though, the article has improved, and this review should pass, preferably with these issues addressed. Tuf-Kat09:56, 17 November 2005 (UTC)[reply]
I'm OK with it. I'd like to see a few of the one-sentence paragraphs addressed, and there's a LOT of text to digest, but it's still up to FA level. I'd recommend passing the review. PacknCanes | say something!18:58, 17 November 2005 (UTC)[reply]
What is the difference between the introduction "Overview" and the "Basic Features" sections? They both seem to be discussing a general overview of the subject (features of a general overview, if you will). Perhaps replace this (ENa) with this (ENa+). Similarly, (EK) with (EK+).
Agreed, much improved. I'd like to add one more suggestion: the second paragraph of "Control of timing" looks like it's basically a glorified see also -- there should be some prose using those terms and explaining what they mean and why they're relevant (and linking to them). Tuf-Kat06:19, 28 November 2005 (UTC)[reply]
I had a look at the "Control of Timing" section. Its largely redundant to the rest of the article. The only real issue that is not redundant is the release of neurotransmitter during the falling phase. That is not really about the action potential so much as it is about the property of calcium channels and tail currents. The part about refractory period is mostly just plain incorrect. The refractory period is primarily derived from sodium channel inactivation, not the undershoot. So I've taken the section out. I'll paste it below in case someone wants to work on it. Synaptidude19:34, 2 December 2005 (UTC)[reply]
Control of timing
Prototypically, an action potential's rising and falling phases are complete in roughly two milliseconds, while the undershoot can last several times longer, depending on the cell. In neurons, the exact length of the roughly two-millisecond delay in the falling phase can have a strong effect on the amount of neurotransmitter released at a synapse. The duration of the overshoot determines a nerve's refractory period and hence the frequency at which it will fire under continuous stimulation. Both of these properties are subject to biological regulation, primarily by acting on potassium channels.
When reading the article from a signal processing POV, there are probably citations for the characteristic impedance of the axons, from Node to Node. The sentences hit all around the concept that there is a similarity between a transmission line and an axon, such as the statement that an action potential's shape is preserved as the pulse propagates down the axon. That is transmission line behavior. --Ancheta Wis00:45, 6 December 2005 (UTC)[reply]
Propagation velocity - size vs. myelin
I think this article would benefit from a bit more information on the mathematics of action potential velocity due to membrane capacitance and axonal resistance, and how these are affected by the diameter and myelination of the axon.
While the "Detailed mechanism" section talk about capacitance, it doesn't mention resistance, which also has a an important role. Comparing the invertebrate and vertebrate methods of increasing action potential velocity is a good way of explaining this topic as it shows the interaction between membrane thickness and axonal diameter (and hence membrane capacitance and axonal resistance). I can add this information, including equations and examples of action potential velocities at different diameters, but thought I check to see what everyone else thinks first... I'm new to this! Hbdgaz16:52, 11 December 2005 (UTC)[reply]
Simplyloic (talk·contribs) has made some substantial edits to the "Underlying mechanism" section. They're much appreciated, but might be a sliver short of featured quality. A few more eyes doing some fact checking and copyediting would be much appreciated. I'll see about merging the new material with what was there previously (see [1]) as well. Cheers, David Iberri (talk) 12:05, 7 April 2006 (UTC)[reply]
hi resting potential section has definetely improved. thanks. But I guess it may contain some more detailed information. for example it may be nice to talk about goldman equation. English is not my mother tongue but in this line "The membrane is said to be polarized.". said to be is unnessesary for me. It is indeed polarised! no discussion or question about it or the usage of the word Polarised is not problematic right? What I would point is the main mechanism that creates the potential is actually membrane selectivity rather than the atpase pumps. these pumps are of importace at rapidly maintaining membrane potential after an action potential. Also the polarity of the membrane proteins add to the membrane resting potential but it is generaly beyond textbook knowledge. keep up the good work :)!neurobio22:52, 6 June 2006 (UTC)[reply]
I agree that the sentence you call attention to seems somewhat out of place, though it may be helpful to briefly introduce layreaders to the concept of membrane polarization. I'm on the fence on this. In either case, it should be rephrased at the very least so as to not sound so out of place.
On the issue of whether ion pumps contribute to the resting potential more or less than membrane permeability to sodium and potassium: The paragraph discussing Na,K-ATPase isn't (IMO) trying to establish its relative importance in generating Vm. It's merely pointing out 1) that ions are differentially distributed across the membrane, 2) Na-K pumps are necessary for generating this distribution, and 3) this distribution is necessary for establishing Vm. The paragraph that follows discusses the importance of the membrane's permeability to potassium in the generation of Vm, and further information is available on the linked article, Resting potential. Cheers, David Iberri (talk) 13:06, 7 June 2006 (UTC)[reply]
Propagation Picture
The traditional current circulation is from the + pole to negative one.
The real current circulation is from negative pole to positive.
The T4 phase of the picture shows a green arrow (circulation) that doesn't respect the rules used in the previous phases.--Somasimple04:57, 8 October 2007 (UTC)[reply]
The main impediment to conduction speed in unmyelinated axons is membrane capacitance. In an electric circuit, the capacity of a capacitor can be decreased by decreasing the cross-sectional area of its plates, or by increasing the distance between plates.
Hi all,
I think that the explanation fails with known facts? If speed seems to be linked with capacitance and if low capacitance promotes speed, why are large fibres (whith large capacitance) quicker than small ones?
There is a contradiction?
--Somasimple05:26, 21 June 2006 (UTC)[reply]
The article doesn't contain a contradiction, although it can perhaps be worded better. You are referring to two distinct and independent physical properties that affect the speed of AP propagation: internal resistance (which is mainly affected by axon diameter) and membrane capacitance (which is mainly affected by myelination status).
AP propagation occurs through the movement of charges. The faster these charges move, the faster the AP propagates. The greater the internal resistance, the more resistance there is to the flow of these charges, and the slower the AP propagates. Internal resistance is a function of axon diameter, with large caliber axons having relatively low resistance compared to small caliber axons. Consequently, large caliber axons will propagate APs faster than small caliber axons, all other things being equal.
Capacitance is the ability to store charge, with increasing values of capacitance indicating a greater ability to store charge. By extension, membrane capacitance is the ability of the membrane to store charged particles. If the membrane sequesters these charges, then they cannot participate in the propagation of the AP. Thus high membrane capacitance results in slower AP propagation, while low capacitance yields rapid propagation. In electrical circuits, a capacitor's capacitance can be decreased in a number of ways: decreasing the cross-sectional diameter of the plates (don't confuse this with the cross-sectional area of the axon), increasing the distance between the plates, or by adding an insulator between the plates. In an axon, the plates are the charges on either side of the membrane, and myelin does the job of increasing the separation between plates and of adding an insulator (fat) between the plates. Thus myelinated axons result in lower membrane capacitance, which increases the rate of AP propagation compared to unmyelinated axons, all other things being equal.
In your question, you seem to think that large fibers have large capacitance. I hope you now understand why this is not necessarily the case, and that axon diameter affects internal resistance, not membrane capacitance. I hope that helps. Let me know if there's anything I or others can clarify. Cheers, David Iberri (talk) 16:20, 21 June 2006 (UTC)[reply]
Hi David,
Hmm, If capacitance is the abillity to store charge then large unmyelinated fibres may get perhaps a lower internal resistance but a larger capacitance. It is a physical rule => diameter increased = more circumference and more charges allowed to stick on the membrane. It is a known fact that the growth curve of speed/diameter is quite a power of 0.6. It shows a slope decrease for large unmyelinated fibres. It is just why nature invented myelinated fibres. So I think that larges fibres (unmyelinated) have larger capacitance than smaller ones. In that case, the statement saying that capacitance is the key issue becomes wrong for unmyelinated fibres because small fibers comparatively are faster.--Somasimple17:05, 21 June 2006 (UTC)[reply]
I'm not familiar with the speed/diameter growth curve you mention. What I can tell you is that the comparisons you're making aren't particularly useful because in each, you're varying more than one property at a time. There was a reason in my earlier post I ended my paragraphs with "all other things being equal". When comparing two things qualitatively as we are doing here, it's important to isolate a single changing variable, holding all others constant. Also, when saying something has larger capacitance (as in your first sentence), it's important to indicate what this is in relation to.
Here are some meaningful comparisons that we can make in looking at the effects on AP propagation of myelination status, membrane capacitance, axon diameter, and internal resistance:
Given two equally myelinated/unmyelinated axons: 1) both axons will have approximately equal membrane capacitance, 2) the axon of greater diameter will have a lower internal resistance, thus 3) in both myelinated and unmyelinated axons, the key factor determining AP propagation speed is axon diameter (ie, internal resistance).
Given two axons of equal diameter: 1) both axons will have the same internal resistance, 2) the axon with greater myelination will have a lower membrane capacitance, thus 3) in axons of equal diameter, the key factor determining AP propagation speed is myelination status (ie, membrane capacitance).
I don't doubt you that axon diameter plays some role in membrane capacitance (that's why I used the phrase "approximately equal membrane capacitance" above). But its contribution to membrane capacitance is vastly smaller than its contribution to internal resistance. --David Iberri (talk) 17:56, 21 June 2006 (UTC)[reply]
Given two equally myelinated/unmyelinated axons...the key factor determining AP propagation speed is axon diameter (ie, internal resistance)
Not sure, because you changed a key factor: the physical resistance of the axon enveloppe. It is well known that unmyelinated fibres bulge during AP propagation. Increasing the thickness of membrane is a way to workaround this "problem".
I did not change a key factor. In the quoted sentence, I clearly referred to the effect of axon diameter on internal resistance. Note that internal resistance is not the resistance of the axonal membrane. I should also say that I am not familiar with the AP bulge of which you speak, and I would be hard-pressed to believe that the lipid bilayer actually thickens during AP propagation. Such an action would presumably require the coordinated action of several enzymes, a biochemical process that would take more time than is practical during AP propagation. --David Iberri (talk) 07:00, 22 June 2006 (UTC)[reply]
Ions are crossing the membrane and are exchanged near it in a thin (virtual) layer of few nanometers. See R MacKinnon. --Somasimple08:15, 22 June 2006 (UTC)[reply]
Agreed. These ions are responsible for the generation of the AP. But as you know, in the myelinated axon segments, no AP is generated; rather the impulse flows passively and in a graded fashion, making internal resistance important. --David Iberri (talk) 18:21, 22 June 2006 (UTC)[reply]
Thirdly, in unmyelinated, we could say that "communication" occurs perpendicularly to the axon membrane. In meylinated, horizontal communication is favored. Could we conclude that Nature used the same means in the two solutions?
Well, If it is not true and we are observing incoming Na+ ions and outgoing K+, does these flux have another orientation? It has been expressed that ions stay in place (do not move horizontally) with marked ions in some experiments. --Somasimple08:15, 22 June 2006 (UTC)[reply]
I think I understand what you are saying. As unmyelinated segments must regenerate the AP at each segment, the impulse has no passive/graded component (such as that present in the AP of a myelinated axon). Thus the relevant charges contributing to an AP in an unmyelinated axon are moving perpendicular to the membrane for the most part. I'm not sure I see what you're trying to get at with the could we conclude that Nature used the same means... question. --David Iberri (talk) 18:21, 22 June 2006 (UTC)[reply]
AP propagation occurs through the movement of charges. I agree totally in unmyelinated fibres since ions are charged particles moving physically. But since we know that they can't travel so far, how do you understand the movement charge within myelinated fibers?--Somasimple04:59, 22 June 2006 (UTC)[reply]
I can't take it since the coulomb's forces falls with the square distance. And you're knowing that myelininated portions are 200 to 2000 time longer than nodes. This simple fact may completely discard such a behaviour since there are tons of ions that are closer than those of the next node. --Somasimple08:15, 22 June 2006 (UTC)[reply]
The analogy to Newton's cradle isn't drawing a correspondence between the cradle's pendula and the axon's nodes of Ranvier. That is, ions aren't leaping from node to node, as you seem to think the article suggests. Rather, the cradle's pendula represent the very closely apposed ions of which you speak; these ions are present throughout the length of the axon's myelinated and unmyelinated segments. When an AP is generated at a node, an electrostatic force is generated by the incoming cations sufficient to repel adjacent cation A a very short distance. Now a hair closer to the next adjacent ion B, a sufficient electrostatic force is generated to repel ion B a short distance. This passive movement of charge occurs many times within the myelinated axon segment until the propagating wave of positivity reaches the next node, whereupon another AP is generated. I hope that clarifies the analogy to Newton's cradle. --David Iberri (talk) 18:21, 22 June 2006 (UTC)[reply]
BTW, you're saying that electrostatic forces are the key engine of propagation (I share the same idea with some variations) then you're telling us that capacitance and resistance are not important. It seems contradictory with the previous statements. --Somasimple08:15, 22 June 2006 (UTC)[reply]
I don't know where I said that capacitance and resistance are not important. Indeed they're very important in considering propagation speed, as can be seen in the determination of the time constant (τ = RC) as Nrets' has explained below. --David Iberri (talk) 18:21, 22 June 2006 (UTC)[reply]
In the figure of propagation, you shows a T4 time with a backward current made impossible by the real nature of AP. Since Ap is a travelling wave then all points exist at the time and thus your left points are ever more positive when you consider the start of the curve. If we get more positive charges on the left, positive ions can't go there.--Somasimple 14:18, 21 June 2006 (UTC)
here is flash anim showing the ingoing positive charges making impossible the left current.
[2]
--Somasimple14:22, 21 June 2006 (UTC)[reply]
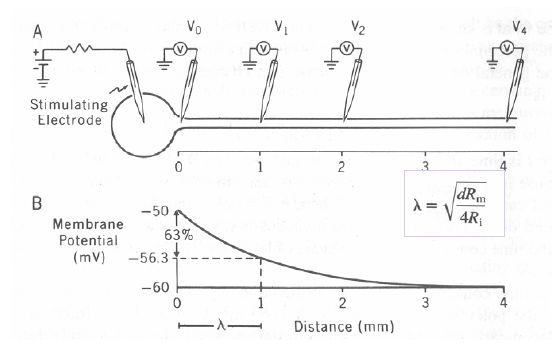
Why do thicker axons conduct faster? My understanding is that conduction velocity will depend on the length constant and time constant of the axon. Thicker axons have a longer length constant since this is = square root of (membrane resistance / axial resistance). Both rm and ra decrease with larger diameters, but ra decreases faster since it is inversely proportional to the area of the cross section of the axon and rm is inversely proportional to the circumference. So fatter axons have a longer length constant, and thus conduct faster. A faster (smaller) time constant will also increase speed of AP propagation. The time constant = Input capacitance x input resistance. For a section of axon, input capacitance = specific membrane capacitance (cm) x circumference of axon; and input resistance = specific membrane resistance (rm) / circumference. Since time constant is = Cinput x Rinput, rearranging the formula you get that time constant = cm x rm. Both cm and rm are properties of the membrane, independent of amount of membrane. Therefore when you have myelin, cm decreases, thus time constant decreases, thus propagation velocity increases. Having a fatter axon does not change the length constant, thus the increase in length constant is what matters. Hope this helps! Nrets15:30, 22 June 2006 (UTC)[reply]
Both rm and ra decrease with larger diameters, but ra decreases faster since it is inversely proportional to the area of the cross section of the axon and rm is inversely proportional to the circumference.
I can't take it since the whole axon "lumen" is not involved. All scientists agree, now that only a thin space of the internal axon is "used" during AP.--Somasimple05:48, 23 June 2006 (UTC)[reply]
Thanks for the reply but no. We started with a legitimate perspective where ions moved accross the membrane and were driven by electrostatic forces. The formulas you brought are pure electrical ones that I cannot use easily with ions that attract and repel.
Ions carry their own forces and of course, more you get, more you may have some speed. The whole is driven, of course, by ions channels which permit these physical ions fluxes.
You have to consider that ions move with water molecules and more they move, more they carry. It is a volume that is never used as a physical mean in AP. It seems also logical to think that in small axons, the membrane resistance (physical property to change shape) is a component that has less importance because ions have few chances to move it. In larger ones, membranes may/is deformed by the amount of carried ions. --Somasimple16:25, 22 June 2006 (UTC)[reply]
No but its not the ions entering a leaving the membrane that determine the propagation velocity, it is the passive cable properties of the axon that do. So when Na+ channels open they will depolarize a segment of membrane, this depolarization will then depolarize the surrounding membrane without the requirement of ions flowing into that next piece of membrane. The length annd time constants will determine how fast the adjacent pieces of membrane will depolarize and how fast voltage gated channels in the next segment will reach threshold and open to regenerate the current. Nrets17:50, 22 June 2006 (UTC)[reply]
it is the passive cable properties of the axon that do.
The passive cable property, again. This analogy was made because our nerves are communication lines but this theory works with electrical means (electrons exchanges) not ionic fluxes (ions moving physically). The major contracdictions with the passive cable theory are numerous:
1/Why the transmission is not instaneous in a 2mm length cable. A normal cable has a amplitude decay over meters but not such a delay. A passive property induces an electronic (via electrons) transmissions and electricy travels at light speed. Under myelin transmission is very much slower.
2/ A light pressure applied over the myelin stops the transmission! Why?
Transmission through an axon is not the same as transmission through a copper cable, true, but it is equivalent of transmission through an ionic solution where the solution is partly insulated via a semi permeable membrane. The effects of the specific membrane resistance and capacitance are to basically cause some of this current to leak out, limiting how far the current can spread down the axon. Both time constants and length constants have been measured empirically and can be predicted by properties such as membrane resistance and membrane capacitance, thus a cable analogy is useful. Measurements of conduction velocities in a wide range of nerve cells have been shown to correlate closely to their calculated resistances and capacitances. Nrets15:45, 23 June 2006 (UTC)[reply]
Thanks, for your reply. Is there a chance to get a reference to the experiements?
What about the response about the delay? Was it found in the experience?
My guess about the delay is that, compared to a real cable the insulation around an axon is much lower so the decay is faster, plus it would be that because of the membrane capacitance, it takes longer for the voltage to change across a distance. But that's just a guess. That being said, conduction velocity in myelinated axons can range from anywhere between 10-100 m/sec. Which is still quite speedy. Some of the original experiments detailing the cable theory of axons are: Hodgkin, A.L. 1954, J.physiol 125, p.221-224; Huxley and Stampfli, (1949), J. Physiol. 108: 315-339. Experiments comparing axon diameter to conduction velocity: Rushton W.A.H. (1951), J. Physiol. 115:101-122; Arbuthnott, Boyd and Kalu (1980), J.Physiol. 308:125-157. If you would like a textbook explanation of the above, I would recommend: Foundations of Cellular Neurophysiology, Johnston and Wu, MIT press, 1995. Hope this helps! Nrets16:54, 23 June 2006 (UTC)[reply]
Thanks for the references but I failed to find the awaited response to my asking. I tink you're talking about these experiments? capacitanceand axon decay These images uncover electrical properties to axon but axon is then in an artificial electrical circuit. My previous asking is a known delay that occurs under meylin during passive transmission.
A movie of this process (50K quicktime animation) shows the influx of sodium as a function of distance along the axon and time. One on the vertical scale indicates the peak of the action potential; the movie makes it clear that the propagation of the impulse occurs much faster (22 µs between node firings for an axon radius of 5 µm) than the time scale of the action potential at a single node (1-2 ms).
I'm not quite sure then I understand what you are asking, the principles underlying passive propagation along the axon are the same with or without myelin. With myelin, the only difference is that Rm is larger and Cm is smaller, leading to a longer legth constant and a faster time constant. From these values you can calculate the rate of change in Vm at a given distance and time from the current source, be that a node with Na+ channels or an artificial current injection. This should explain the delay between nodes. The first 2 refs should address this, but also look at the textbook since it will summarize years of work into a cohesive picture. Nrets18:44, 24 June 2006 (UTC)[reply]
Imagine that Action potential is a solar car (ions are mimicking the sun). I can understand that when the sun is lighting my car, my car can travel.
But in the original theory, they are simply saying that if the car enters a long tunnel, the car goes faster. What a smart car! But Nature doesn't make miracles. Nature used a different mean as I thought.
You can't argue with experimental data that shows that myelinated axons conduct faster than non-myelinated ones. Did you look at: Huxley and Stampfli, (1949), J. Physiol. 108: 315-339 ?? Nrets14:21, 27 June 2006 (UTC)[reply]
Nrets, I do not deny that myelinated axons transmit faster an AP but only the theory behind this transmission. You say it is passive and it is quite true but since the speed is increased and because you insist that capacitance and resistance persist, and because AP amplitude decays, and because no ions are injected under the myelin then the passive transmission discards the theory of Huxley and Hodgkin. You can't have a passive transmission with passive 'brakes' (C and R) and have at the same time, a speed increased. It doesn't work like that. It is impossible.--Somasimple14:38, 27 June 2006 (UTC)[reply]
I still don't understand what you are confused about. Basically, if you can calculate the various membrane parameters (Cm, Rm) as well as the geometry of the axon, then you can accurately predict conduction velocity. Plain and simple. How then does this make the theory incorrect? Hodgkin and Huxley specifically modelled the changes in conductance when you have actual voltage gated channels in the membrane, they based their initial theory on experiments on a squid axon which has no myelin. Nrets17:43, 27 June 2006 (UTC)[reply]
Also, someone just added this link to the article and it does a pretty thorough job describing AP propagation and generation. Hope this helps. Nrets19:02, 27 June 2006 (UTC)[reply]
Nrets, you do not listen to me. An AP is a travelling wave. If its speed is v1 at node and v2 under myelin (with v2>v1) AND because it is travelling wave AND thus a continuous phenomenon, every point of the curve MUST be dilated while changing medium. IT IS NOT WHAT SCIENTISTS RECORDED. THE SHAPE OF THE AP IS PRESERVED. CONCLUSION => the shape is not really transmitted. The wave is not travelling under the myelin.
I have already read the Benazilla's stuff. A great scientist but too electrical.--Somasimple04:40, 28 June 2006 (UTC)[reply]
If you sample the wave with the same timeline (like HH did it), a wave that goes faster under myelin will be mandatory shrunk. I will provide some pictures/movies on my site.--Somasimple05:00, 28 June 2006 (UTC)[reply]
Nrets, I put again the link of registered events under myelin => under myelin. You are seeing a graph of AP transmitted with time and distance. The shape is preserved. The shape continues to grow at previous places and begins to grow at further distances. If the wave exists at many places at the same time, the shape didn't move. We are seeing it further because it grows at previous places.--Somasimple05:45, 28 June 2006 (UTC)[reply]
It's not clear from the movie you show, what any of those axes are (time, distance, volume, voltage?) so I can't comment on that, but I did look at your website and I think your fundamental misunderstanding may lie in your analogy: "Imagine that Action potential is a solar car (ions are mimicking the sun). I can understand that when the sun is lighting my car, my car can travel. But In the original theory, they are simply saying that if the car enters a long tunnel, the car goes faster. What a smart car! But Nature doesn't make miracles. Nature used a different mean as I thought." This is not a good analogy, for the car is only being propelled by energy it receives. During an action potential propagation is active where there is no myelin, but is also passively propagating all the time. Passive propagation is facilitated in a myelinated axon even if the current does not regenerate while it travels under myelin. The best way to understand passive propagation is to study subthreshold signals that do not activate voltage gated channels. Once again, the membrane properties CHANGE between myelinated sections of the axon and non-myelinated sections of the axon therefore the efficiency of propagation changes. This is sufficient to explain experimental observations, and you do not need to invoke active changes in axonal volume to expalin them. If you had read the Benzanilla tutorial carefully, you should see that this is the case. Also, I would recommend textbooks like "From Neuron to Brain" or "Fundamentals of Neurophysiology" or even "Principles of Neuroscience" (but this isn't quite as good) for more detailed explanations. The online text that you quote from in your site and post a figure from is a molecular biology text book, and really does not do a good job at explaining AP propagation. Hope this helps and good luck! Nrets14:50, 28 June 2006 (UTC)[reply]
Nrets, I took the same references than you. I took this example because it seems poor, intentionally. Imagine that AP is a train and as an observer, you're taking pictures of the running train. If the rate of taking pictures is fixed then you can take 100 pictures if the speed of the train is v1 and only 50 if the speed v2 is 2 times higher than the first train. If you put the taken pictures on a table, you've got a long train and a short one: It depends of the speed of the trains. ALL EXPERIMENTS RECORD A SAME LENGHT OF AP. THE SAME LENGHT, WITHOUT OR WITH MYELIN. Just use your logic and your brain, forget for a while, what you've learnt. How could you explain it? Good luck!--Somasimple15:35, 28 June 2006 (UTC)[reply]
Soma, it makes more sense to measure duration of an AP rather than length (which is not very intuitive). If you recorded a single AP in a myelinated axon vs. a non-myelinated axon, you would see that in the myelinated axon the rise and decay times of the AP would be faster. Likewise if you record at 2 points along the same axon, one at the site of a subthreshold current injection and one distant to it. At the distant site in the myelinated axon you would detect that the voltage has decreased less at a given point in time after the current injection than in an unmyelinated axon. This makes perfect sense both intuitively and theoretically. So stop acting so condescendingly, I'm only trying to help. Nrets17:56, 28 June 2006 (UTC)[reply]
Nrets, the length term was employed as duration, of course. Do you mean that my example is one more time, useless? Or not logical? What about the barrier beetween the node and the myelin? What is the phenomenon able to "elongate" the curve? What about water injected at the site/node? temperature? membrane deformation? You're constantly replying with 60 years olds papers! Did the research stopped in 1952? You are replying with evasive responses, it doesn't help.--Somasimple19:29, 28 June 2006 (UTC)[reply]
No, the reaserch moved on. You are the one who is dismissing everything I say. I'd be very interested to see some references to studies which show direct evidence that AP conduction velocity cannot be predicted by passive and active membrane properties. I'm not saying this isn't possible, I just don't know of any. Nrets19:47, 28 June 2006 (UTC)[reply]
You wrote:"I'd be very interested to see some references to studies which show direct evidence that AP conduction velocity cannot be predicted by passive and active membrane properties.". Nrets, since AP occurs in an axon nobody can ;-) You can follow my foolish discussion on my site. You're welcome! ps: I have clues and proves.--Somasimple04:01, 29 June 2006 (UTC)[reply]
Agreed 100%. I've hated that sentence for quite some time. ;-) It's especially peculiar in the introduction, as the article never expounds on this idea. --David Iberri (talk) 07:02, 22 June 2006 (UTC)[reply]
I took a quick look in the research literature and found some articles that use the term "neural code".
Spike firing allometry in avian intrapulmonary chemoreceptors: matching neural code to body size "In general, neural information may be `rate coded,' with average spike rate over a time period encoding stimulus intensity, or `time coded,' with the occurrence of a single spike (or spike burst) encoding the occurrence of a rapid stimulus transition.....Rate and time codes are not mutually exclusive: `partially adapting' sensory neurons are common, and have both tonic (rate coded) and phasic (time coded) discharge responses"
Variability and information in a neural code of the cat lateral geniculate nucleus - "neural code" seems to be used to mean the information carried by action potentials passing from one brain region to another. The is no on "neural code", rather each pair of connected brain regions is suspected of having a specialized "code" that is particularly efficient for dealing with the type of information being sent.
Deciphering a neural code for vision - this article seems to use "neural code" to mean how the information available in visual sensory cell activity is converted into a pattern of action potentials in the optic nerve....the implication being that the action potential pattern must later be "decoded" in order for downstream brain regions to obtain the original sensory information.
My conclusions; 1) It is a matter of on-going research to determine how information is sent between brain regions in the form of sets of action potentials....there are different types of "neural codes" that are used by different neurons. 2) Action potentials are important in skeletal muscle cells where "neural codes" are irrelevant. 3) It does not seem reasonable to introduce "neural code" into the introductory section of the article.
Possible alternative. Replace, "Action potentials are an essential carrier of the neural code," with something like, "Action potentials are rapidly transmitted electrical signals that complement slower chemical signals. --JWSchmidt15:23, 22 June 2006 (UTC)[reply]
They don't realy complement slower chemical signals (which really are quite fast) rather they work in series with them. Maybe we should say: "Action potentials are rapidly transmitted electrical signals used by brain cells to convey neural information." Nrets18:23, 22 June 2006 (UTC)[reply]
plant expansion in the article or removal from intro
The introduction has one whole paragraph on the electrical signals in plants but nothing in the main text. While this is intereting and relevant this text should be moved from the introduction to the main text. The introduction is supposed to be a summary, not a place for new information. Alternatively a much larger section on electrical signals in plants should be incorporated into the article.
Personally i think it would be enough to remove the text from the introduction. This topic should be mentioned in the main text but refer to a separate article. David D.(Talk)15:51, 23 June 2006 (UTC)[reply]
There are lots of things that make action potential firing more or less likely, such as neuromodulators, tonic inhibition, channel density, etc. I think if you do mention LTP/LTD it should be in the context of things that alter neuronal excitability. Nrets15:11, 1 October 2006 (UTC)[reply]
Questions
Do you suppose it's worth saying anything about the fact that Hodgkin & Huxley got
the Nobel prize for figuring out how action potentials work?
It seems odd that the idealized sketch of an action potential has a rising phase that is so slow compared with its falling phase. It also seems odd for this figure to have such a salient line labeled "threshold"--as if there is such a thing as "the" threshold for triggering a spike.
This statement
"the threshold of an action potential is not the voltage at which sodium current becomes significant; it is the point where it exceeds the potassium current."
is true only for a membrane action potential (i.e. only for a nonpropagating spike
in an isolated patch of membrane). It is not true for action potentials that arise
at a "trigger zone" in a physically extended structure, such as a neuron (or even
an axon).
"An action potential is a rapid swing in the polarity of the voltage from negative to positive and back, the entire cycle lasting a few milliseconds. Each cycle—and therefore each action potential—has a rising phase, a falling phase, and finally an undershoot"
Except for those cells that have broad action potentials, e.g. cardiac muscle.
The introduction now mentions the fact that there are different types of action potentials; this article describes the "typical" action potential of axons.
The following statement is meaningless:
"The natural tendency of sodium and potassium ions is to diffuse across their electrochemical gradients to attempt to reach their respective equilibrium potentials"
That sentence has been changed.
If "the resting cell membrane is approximately 100 times more permeable to potassium than to sodium, so that more potassium diffuses out of the cell than sodium diffuses in", then how can a pump with 3:2 stoichiometry maintain the cell in steady state?
Because the leak conductance is very small. So the overall change in ion concentration inside the entire volume of the cell is very small. This is also the case during an action potential. Nrets18:40, 30 October 2006 (UTC)[reply]
"the dominant outward leak of potassium ions produces a hyperpolarizing current that establishes the cell's resting potential of roughly -70 mV."
A cell at rest has a net hyperpolarizing transmembrane current?
Yes, otherwise how do you think you acheive a resting membrane potential. The membrane potential is a steady state where net inward and outward currents are equal, thus you have a resting K+ connductance and a resting Na+ conductance.
If "inward and outward currents are equal," net transmembrane current is 0, not hyperpolarizing.
"Establishment of a membrane potential of around +40 mV closes the voltage-sensitive inactivation gates of the sodium channels, which are sensitive to the now-positive membrane potential gradient, preventing further influx of sodium. While this occurs, the voltage-sensitive activation gates on the voltage-gated potassium channels begin to open."
Which implies the following totally incorrect conclusions:
1. sodium current turns off at, or shortly after, the peak of the spike
According to the H&H model it does. After the Na+ channels inactivate, the Na+ conductance starts to decrease.
The statement "Establishment of a membrane potential of around +40 mV closes the voltage-sensitive inactivation gates of the sodium channels . . . preventing further influx of sodium" is false.
A. "According to the H&H model it does." Not so. Sodium current does not turn off at the peak of the spike, or shortly after the peak. In fact, not only does it continue to increase for quite some time after the peak of the spike, (1) the maximum sodium current (which occurs during the falling phase of the spike) is almost twice as big as the value at the peak of the spike, and (2) about 80% of the sodium influx during a spike occurs _after_ the peak.
B. "After the Na+ channels inactivate, the Na+ conductance starts to decrease." Again not so. Inactivated channels are closed channels. Closed channels have zero conductance. Their conductance does not "start to decrease after they inactivate." And in the HH model, sodium conductance continues to increase for a bit after the peak of the spike.
2. most of the sodium influx during a spike occurs before the depolarized peak; very little enters after the peak
I don't think the sentence implies this at all. Obviously if the decay phase of the Na+ conductance is slower than the rising phase, then yes, you will get more net Na+ influx during the falling phase of the Na+ conductance. But I don't see where it is implied that more Na+ enters the cell during the rising than falling phase.
The statement is that "Establishment of a membrane potential of around +40 mV closes the voltage-sensitive inactivation gates of the sodium channels . . . preventing further influx of sodium"? This is a clear assertion that the sodium channels close and no more sodium influx can occur. It does not say that some channels remain open.
3. sodium current dwarfs potassium current during the rising phase of the spike
Again, according to the H&H model during the initial phase of the AP Na+ conductances are much larger than K+ conductances, which have a slightly delayed opening (thus the name delayed rectifier).
The delay is only slight. At the point of maximum slope on the rising phase of the spike, potassium current is already about 20% as large as sodium current. And it catches up quickly--at the peak of the spike, the currents are nearly equal.
[Action potentials] "are often modeled using hypothetical cells containing only two transmembrane ion channels (a voltage-gated sodium channel and a non-voltage-gated potassium channel)."
Citation please. Somewhere or other in this article, one might expect to see the
Hodgkin-Huxley model mentioned, too, if merely as a counterexample.
The discussion of IV curves lacks clarity.
"The main impediment to conduction speed in unmyelinated axons is membrane capacitance."
Whatever happened to cytoplasmic resistivity? By the way, cytoplasmic resistivity accounts for the effect of axon diameter on conduction velocity.
Yes, but in the case of myelination, membrane capacitance is more important. I wouldn't object to the mention internal resistance in this sentence if you feel strongly about it.
Membrane capacitance would have no effect on conduction velocity if it were not for cytoplasmic resistivity. That's a consequence of the laws of physics, not a matter about which feelings have any relevance.
[Myelination] "allows the rapid (essentially instantaneous) conduction of ions through a myelinated segment of axon"
Not so. Ionic mobility is very slow--far slower than the action potential.
True it should read conduction of current.
Bear et al., Kandel's books, and Purvis et al. are very weak on this stuff. Aidley (The Physiology of Excitable Cells) is a far superior text, to name just one.
Why don't you add it to the list of refs?
In general I think the anonymous editor gives great suggestions, but if he does make changes to the article, please make sure to keep it in more accessible language, as this is a general use encyclopedia, rather than a specialized textbook. Nrets18:33, 30 October 2006 (UTC)[reply]
Possible error
I could be wrong, but isn't this passage incorrect?
An action potential is a rapid swing in the polarity of the voltage from positive to negative and back, the entire cycle lasting a few milliseconds.
Maybe I'm just confused about action potentials, but I thought that the resting potential voltage was negative, then when the action potential came along, there is depolarisation to a positive value, then repolarisation back to negative. So surely it should be:
An action potential is a rapid swing in the polarity of the voltage from negative to positive and back, the entire cycle lasting a few milliseconds.
It all depends on your point of reference. In modern nomenclature, the resting potential of a typical neuron (inside relative to outside) is negative (~60-70mV). However, in the original J. Physiology papers by H-H, they use the opposite reference point (outside relative to inside) hence the resting potential starts positive and the AP causes a transient drop in the potential. This is probably where the confusion comes in (It gets new 1st time readers of those papers every time...)
I thought that the original HH articles reversed the direction of current flow, but the resting voltage was always negative and AP was always positive. Nrets17:37, 13 February 2007 (UTC)[reply]
[3] This article seems to suggest that it is sound waves, not electrical impulses which are propagating. Can anyone confirm? If so, much of this article would seem to now be incorrect.
Cariaso22:54, 6 March 2007 (UTC)[reply]
That research is far from established dogma. Until the hypothesis is independently confirmed, there's no need to update this article. --David Iberri (talk) 23:15, 6 March 2007 (UTC)[reply]
Having read the paper, the summary article spins the original paper more than the original authors do...which is a lot. What can be fairly drawn from the original paper is that non-specific anesthetics may impart anesthetic affect by altering the ability of the membrane to move in response to electrical stimulation. This could affect the ability of proteins in the membrane to transition from one state to another (thus leading to anesthesia), though this has yet to be demonstrated. The original authors did try to spin their data to argue that piezoelectric pulses underlie nerve conduction, but this is trivially disproven by even the most casual glance at the literature (e.g. noting channelopathies and knockout animals). They're lucky I wasn't one of their reviewers. So, we can safely conclude that the Action Potential article is fine. --Dpryan21:38, 7 March 2007 (UTC)[reply]
plant a/p
can someone provide a reference for plant actin potential. i've never come across this, i sounds facinating! oh yeah and because that section is unreferenced as well i guess.--Dylan210610:50, 14 May 2007 (UTC)[reply]
The "RC circuit" picture - wrong?
It seems to me that part B of the image RC_membrane_circuit.jpg is wrong. It should be like for example here:
http://biology.unm.edu/toolson/b435/equivcirc.html, the batteries in series with
the resistors (channels).
Yes, you are both right. This is my diagram, and it is, in fact incorrect. The batteries should, in fact, be in series with the resistors. I've been meaning to fix it for a while, and just did not get around to it. I'll so it soon. Synaptidude
The traditional current circulation is from the + pole to negative one. The real current circulation is from negative pole to positive. The T4 phase of the picture shows a green arrow (circulation) that doesn't respect the rules used in the previous phases.--Somasimple06:50, 9 October 2007 (UTC)[reply]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}