
Sonic hedgehog protein
Template:PBB Sonic hedgehog homolog (SHH) is one of three proteins in the mammalian signaling pathway family called hedgehog, the others being desert hedgehog (DHH) and Indian hedgehog (IHH). SHH is the best studied ligand of the hedgehog signaling pathway. It plays a key role in regulating vertebrate organogenesis, such as in the growth of digits on limbs and organization of the brain. Sonic hedgehog is the best established example of a morphogen as defined by Lewis Wolpert's French flag model—a molecule that diffuses to form a concentration gradient and has different effects on the cells of the developing embryo depending on its concentration. SHH remains important in the adult. It controls cell division of adult stem cells and has been implicated in development of some cancers.
Discovery
The hedgehog gene (hh) was first identified in the classic Heidelberg screens of Eric Wieschaus and Christiane Nusslein-Volhard, as published in 1978. These screens, which led to their winning the Nobel Prize in 1995 along with developmental geneticist Edward B. Lewis, identified genes that control the segmentation pattern of Drosophila melanogaster (fruit fly) embryos. The hh loss of function mutant phenotype causes the embryos to be covered with denticles (small pointy projections), resembling a hedgehog.
Investigations aimed at finding a hedgehog equivalent in mammals revealed three homologous genes. The first two discovered, desert hedgehog and Indian hedgehog, were named for species of hedgehogs, while sonic hedgehog was named after Sega's video game character Sonic the Hedgehog.[1] In zebrafish, the orthologues of the three mammalian hh genes are: shh a,[2] shh b,[3] (formerly described as tiggywinkle hedgehog named for Mrs. Tiggy-Winkle, a character from Beatrix Potter's books for children), and indian hedgehog b[4] (formerly described as echidna hedgehog, named for the spiny anteater, though this may have also been a playful reference to Knuckles the Echidna, another character from the Sonic the Hedgehog series of video games).
Function
Of the hh homologues, shh has been found to have the most critical roles in development, acting as a morphogen involved in patterning many systems, including the limb[5] and midline structures in the brain,[6] spinal cord,[7] the thalamus by the zona limitans intrathalamica[8] and the teeth.[9] Mutations in the human sonic hedgehog gene, SHH, cause holoprosencephaly type 3 (HPE3) as a result of the loss of the ventral midline. Sonic hedgehog is secreted at the zone of polarizing activity (ZPA), which is located on posterior side of a limb bud in an embryo. The sonic hedgehog transcription pathway has also been linked to the formation of specific kinds of cancerous tumours.
More recently, sonic hedgehog has also been shown to act as an axonal guidance cue. It has been demonstrated that Shh attracts commissural axons at the ventral midline of the developing spinal cord.[10] Specifically, Shh attracts retinal ganglion cell (RGC) axons at low concentrations and repels them at higher concentrations.[11] The absence (non-expression) of Shh has been shown to control the growth of nascent hind limbs in cetaceans[12] (whales and dolphins).
Patterning of the central nervous system
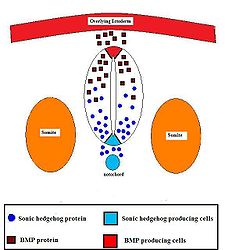
The Sonic hedgehog (Shh) signaling molecule assumes various roles in patterning the central nervous system (CNS) during vertebrate development. One of the most characterized functions of Shh is its role in the induction of the floor plate and diverse ventral cell types within the neural tube.[13] The notochord, a structure derived from the axial mesoderm, produces Shh which travels extracellularly to the ventral region of the neural tube and instructs those cells to form the floor plate.[14]
The neural tube itself is the initial groundwork of the vertebrate CNS, and the floor plate is a specialized structure located at the ventral midpoint of the neural tube. Evidence supporting the notochord as the signaling center comes from studies in which a second notochord is implanted near a neural tube in vivo, leading to the formation of an ectopic floor plate within the neural tube.[15]
-
 Shh and BMP gradients in the vertebrate neural tube
Shh and BMP gradients in the vertebrate neural tube -
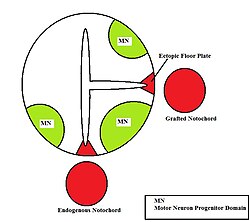 Ectopic floor plate formation
Ectopic floor plate formation -
 Ventral neural domains in neural tube
Ventral neural domains in neural tube



Evidence for Shh as the signaling molecule emanating from the notochord comes from studies involving mutant mice. These mice, devoid of Shh signaling activity but not a notochord structure, fail to develop the floor plate.[16] Shh exerts its effects in a concentration-dependent manner,[17] so that a high concentration of Shh result in a local inhibition of cellular proliferation.[18] This inhibition causes the floor plate to become thin compared to the lateral regions of the neural tube. Lower concentrations of Shh results in cellular proliferation and induction of various ventral neural cell types.[19] Once the floor plate is established, cells residing in this region will subsequently express Shh themselves.[18] generating a concentration gradient within the neural tube. (Although there is no direct evidence of a Shh gradient, there is indirect evidence via the visualization of Patched expression throughout the ventral neural tube.[20] The gradient of Shh present within the neural tube establishes several domains of interneuron and motor neuron progenitors[18] as well as oligodendrocyte precursors.[21] Evidence for the role of Shh in motor neuron induction comes from experimental depletion of Shh activity using protein interfering antibodies in vivo, resulting in the absence of differentiated motor neurons.[19]
The different progenitor domains contained by the ventral neural tube are established by “communication” between different classes of homeobox transcription factors. These transcription factors are grouped into class I and Class II genes (depending on their response to Shh activity), and are composed of members from the Pax, Nkx, Dbx, and Irx families.[18] Class I genes and downregulated in response to high Shh concentration (active in dorsal neural tube), and Class II genes are upregulated in response to high Shh concentration (active in ventral neural tube).[22] Each of the two classes of transcription factors are able to repress expression of the other, a mutual antagonism that is able to create regional domains.
It is important to note that Shh is not the only signaling molecule exerting an effect on the developing neural tube. Many other molecules, pathways and mechanisms are active (ex. RA, FGF, BMP); and complex interactions between Shh and other molecules is possible.
Morphogenetic Activity
The concentration and time dependent cell fate determining activity of Sonic Hedgehog (Shh) in the ventral neural tube makes it a prime example of a morphogen. In vertebrates, Shh signaling in the ventral portion of the neural tube is most notably responsible for the induction of floor plate cells and motor neurons.[23]
Shh eminates from the notochord and ventral floor plate of the developing neural tube to create a concentration gradient that spans the dorso-ventral axis. [24] Higher concentrations of the Shh ligand are found in the most ventral aspects of the neural tube and notochord, while lower concentrations are found in the more dorsal regions of the neural tube.[24] The Shh concentration gradient has been visualized in the neural tube of mice engineered to express a Shh::GFP fusion protein to show this graded distribution of Shh during the time of ventral neural tube patterning. [25]

It is thought that the Shh gradient works to elicit multiple different cell fates by a concentration and time dependent mechanism that induces a variety of transcripton factors in the ventral progenitor cells.[24][25]Each of the ventral progenitor domains expresses a highly individualized combination of transcription factors: Nkx2.2, Olig2, Nkx6.1, Nkx 6.2, Dbx1, Dbx2, Irx3, Pax6, and Pax7, that is regulated by the Shh gradient. These transcription factors are induced sequentially along the Shh concentration gradient with respect to the amount and time of exposure to Shh ligand.[24] As each population of progenitor cells responds to the different levels of Shh protein, they begin to express a unique combination of transcription factors that leads to neuronal cell fate differentiation. This Shh induced differential gene expression creates sharp boundries between the discrete domains of transcription factor expression which ultimately patterns the ventral neural tube. [24]
The spacial and temporal aspect of the progressive induction of genes and cell fates in the ventral neural tube is illustrated by the expression domains of two of the most well characterized transcription factors Olig2, and Nkx2.2. [24] Early in development the cells at the ventral midline have only been exposed to a low concentration of Shh for a relatively short time and express the transcription factor Olig2. [24] The expression of Olig2 rapidly expands in a dorsal direction concomitantly with the continous dorsal extension of the Shh gradient over time. [24] However, as the morphogenetic front of Shh ligand moves and begins to grow more concentrated, cells that are exposed to higher levels of the ligand respond by switching off Olig2 and turning on Nkx2.2. [24] Thus, Creating a sharp boundry between the cells expressing the transcription factor Nkx2.2 ventral to the cells expressing Olig2. It is in this way that each of domains of the six progenitor cell populations are thought to be successively patterned throughout the neural tube by the Shh concentration gradient. [24]
Processing

SHH undergoes a series of processing steps before it is secreted from the cell. Newly synthesised SHH weighs 45 kDa and is referred to as the preproprotein. As a secreted protein it contains a short signal sequence at its N-terminus, which is recognised by the signal recognition particle during the translocation into the endoplasmic reticulum (ER), the first step in protein secretion. Once translocation is complete, the signal sequence is removed by signal peptidase in the ER. There SHH undergoes autoprocessing to generate a 20 kDa N-terminal signaling domain (SHH-N) and a 25 kDa C-terminal domain with no known signaling role.[26] The cleavage is catalysed by a protease within the C-terminal domain. During the reaction, a cholesterol molecule is added to the C-terminus of SHH-N.[27] Thus the C-terminal domain acts as an intein and a cholesterol transferase. Another hydrophobic moiety, a palmitate, is added to the alpha-amine of N-terminal cysteine of SHH-N. This modification is required for efficient signaling, resulting in 30-fold increase in potency over the non-palmitylated form.[28]
Robotnikinin
A potential inhibitor of the Hedgehog signaling pathway has been found and dubbed 'Robotnikinin', in honor of Sonic the Hedgehog's nemesis, Dr. Ivo "Eggman" Robotnik.[29]
Criticism of the name
Some clinicians and scientists criticize giving genes frivolous, whimsical, or quirky names, calling it inappropriate that patients with "a serious illness or disability are told that they or their child have a mutation in a gene such as Sonic hedgehog."[30][31]
See also
- Zbtb7, a gene which was originally named "Pokemon"
- Pikachurin, a retinal protein named after Pikachu
- Cyclopia
References
- ^ Anwood, Robert (2007-09-06). Emus Can't Walk Backwards. Ebury Press. pp. 113–114. ISBN 9780091921514.
- ^ "Zebrafish shha". zfin.org.
{{cite web}}: Cite has empty unknown parameter:|coauthors=(help) - ^ "Zebrafish shhb". zfin.org.
{{cite web}}: Cite has empty unknown parameter:|coauthors=(help) - ^ "Zebrafish ihhb". zfin.org.
{{cite web}}: Cite has empty unknown parameter:|coauthors=(help) - ^ Currie P.D. and Ingham P.W. (1996). "Induction of a specific muscle cell type by a hedgehog-like protein in zebrafish". Nature. 382 (6590): 452–5. doi:10.1038/382452a0. PMID 8684485.
- ^ Herzog W, Zeng X, Lele Z, Sonntag C, Ting JW, Chang CY, Hammerschmidt M (2003). "Adenohypophysis formation in the zebrafish and its dependence on sonic hedgehog". Dev. Biol. 254 (1): 36–49. doi:10.1016/S0012-1606(02)00124-0. PMID 12606280.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Lewis KE, Eisen JS (2001). "Hedgehog signaling is required for primary motoneuron induction in zebrafish". Development. 128 (18): 3485–95. PMID 11566854.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ Scholpp S, Wolf O, Brand M, Lumsden A (2006). "Hedgehog signalling from the zona limitans intrathalamica orchestrates patterning of the zebrafish diencephalon". Development. 133 (5): 855–64. doi:10.1242/dev.02248. PMID 16452095.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Dassule HR, Lewis P, Bei M, Maas R, McMahon AP (2000). "Sonic hedgehog regulates growth and morphogenesis of the tooth" (PDF). Development. 127 (22): 4775–85. PMID 11044393.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Charron F, Stein E, Jeong J, McMahon AP, Tessier-Lavigne M (2003). "The morphogen sonic hedgehog is an axonal chemoattractant that collaborates with netrin-1 in midline axon guidance". Cell. 113 (1): 11–23. doi:10.1016/S0092-8674(03)00199-5. PMID 12679031.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Kolpak A, Zhang J, Bao ZZ (2005). "Sonic hedgehog has a dual effect on the growth of retinal ganglion axons depending on its concentration". J. Neurosci. 25 (13): 3432–41. doi:10.1523/JNEUROSCI.4938-04.2005. PMC 1564194. PMID 15800198.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Thewissen JG, Cohn MJ, Stevens LS, Bajpai S, Heyning J, Horton WE (2006). "Developmental basis for hind-limb loss in dolphins and origin of the cetacean bodyplan". Proc. Natl. Acad. Sci. U.S.A. 103 (22): 8414–8. doi:10.1073/pnas.0602920103. PMC 1482506. PMID 16717186.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Litingtung Y, Chiang C (2000). "Control of Shh activity and signaling in the neural tube". Dev. Dyn. 219 (2): 143–54. doi:10.1002/1097-0177(2000)9999:9999<::AID-DVDY1050>3.0.CO;2-Q. PMID 11002335.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ Placzek M (1995). "The role of the notochord and floor plate in inductive interactions". Curr. Opin. Genet. Dev. 5 (4): 499–506. PMID 7580143.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ van Straaten HW, Hekking JW, Thors F, Wiertz-Hoessels EL, Drukker J (1985). "Induction of an additional floor plate in the neural tube". Acta Morphol Neerl Scand. 23 (2): 91–7. PMID 3834777.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA (1996). "Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function". Nature. 383 (6599): 407–13. doi:10.1038/383407a0. PMID 8837770.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Placzek M, Tessier-Lavigne M, Yamada T, Jessell T, Dodd J (1990). "Mesodermal control of neural cell identity: floor plate induction by the notochord". Science. 250 (4983): 985–8. PMID 2237443.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ a b c d Wilson L, Maden M (2005). "The mechanisms of dorsoventral patterning in the vertebrate neural tube". Dev. Biol. 282 (1): 1–13. doi:10.1016/j.ydbio.2005.02.027. PMID 15936325.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ a b Ericson J, Morton S, Kawakami A, Roelink H, Jessell TM (1996). "Two critical periods of Sonic Hedgehog signaling required for the specification of motor neuron identity". Cell. 87 (4): 661–73. PMID 8929535.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Marigo V, Tabin CJ (1996). "Regulation of patched by sonic hedgehog in the developing neural tube". Proc. Natl. Acad. Sci. U.S.A. 93 (18): 9346–51. PMC 38430. PMID 8790332.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ Davies JE, Miller RH (2001). "Local sonic hedgehog signaling regulates oligodendrocyte precursor appearance in multiple ventricular zone domains in the chick metencephalon". Dev. Biol. 233 (2): 513–25. doi:10.1006/dbio.2001.0224. PMID 11336511.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ Briscoe J, Sussel L, Serup P, Hartigan-O'Connor D, Jessell TM, Rubenstein JL, Ericson J (1999). "Homeobox gene Nkx2.2 and specification of neuronal identity by graded Sonic hedgehog signalling". Nature. 398 (6728): 622–7. doi:10.1038/19315. PMID 10217145.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Roelink H, Porter JA, Chiang C, Tanabe Y, Chang DT, Beachy PA, Jessel TM (1995). "Floor Plate and Motor Neuron Induction by Different Concentrations of the Amino-Terminal Cleavage Product of Sonic Hedgehog Autoproteolysis". Cell. 81: 445–455. doi:10.1016/0092-8674(95)90397-6.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ a b c d e f g h i j Ribes V, Briscoe J (2009). "Establishing and interpreting Graded Sonic Hedgehog during Vertebrate Neural Tube Patterning: The Role of Negative Feedback". Cold Spring Harb Perspect Biol. 1(2): a002014. doi:10.1101/cshperspect.a002014.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ a b Chamberlain CE, Jeong J, Guo C, Allen BL, McMahon AP (2008). "Notochord-derived Shh concentrates in close association with the apically positioned basal body in neural target cells and forms a dynamic gradient during neural patterning". Development. 135(6): 1097–1106. doi:10.1242/dev.013086.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) - ^ Bumcrot DA, Takada R and McMahon AP (1 April 1995). "Proteolytic processing yields two secreted forms of sonic hedgehog". Mol Cell Biol. 15 (4): 2294–2303. PMC 230457. PMID 7891723.
- ^ Porter JA, Young KE and Beachy PA (1996). "Cholesterol modification of hedgehog signaling proteins in animal development". Science. 274 (5285): 255–259. doi:10.1126/science.274.5285.255. PMID 8824192.
- ^ Pepinsky RB, Zeng C, Wen D, Rayhorn P, Baker DP, Williams KP, Bixler SA, Ambrose CM, Garber EA, Miatkowski K; et al. (1998). "Identification of a palmitic acid-modified form of human Sonic hedgehog". J Biol Chem. 273 (22): 14037–14045. doi:10.1074/jbc.273.22.14037. PMID 9593755.
{{cite journal}}: Explicit use of et al. in:|author=(help)CS1 maint: multiple names: authors list (link) CS1 maint: unflagged free DOI (link) - ^ "Robotnikinin takes on Sonic hedgehog - robotnikinin, Sonic hedgehog - Australian Life Scientist". www.lifescientist.com.au. Retrieved 2009-09-21.
- ^ Maclean K (2006). "Humour of gene names lost in translation to patients". Nature. 439 (7074): 266. doi:10.1038/439266d. PMID 16421543.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ Cohen MM (2006). "Problems in the naming of genes". Am. J. Med. Genet. A. 140 (13): 1483–4. doi:10.1002/ajmg.a.31264. PMID 16718675.
{{cite journal}}: Unknown parameter|month=ignored (help)
Further reading
- Dorus S, Anderson JR, Vallender EJ; et al. (2006). "Sonic Hedgehog, a key development gene, experienced intensified molecular evolution in primates". Hum. Mol. Genet. 15 (13): 2031–7. doi:10.1093/hmg/ddl123. PMID 16687440.
{{cite journal}}: Explicit use of et al. in:|author=(help)CS1 maint: multiple names: authors list (link) - Gilbert, Scott F. (2000). Developmental biology (6th ed.). Sunderland, Mass: Sinauer Associates. ISBN 0-87893-243-7.
- Kim J, Kim P, Hui CC (2001). "The VACTERL association: lessons from the Sonic hedgehog pathway". Clin. Genet. 59 (5): 306–15. doi:10.1034/j.1399-0004.2001.590503.x. PMID 11359461.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Morton JP, Lewis BC (2007). "Shh signaling and pancreatic cancer: implications for therapy?". Cell Cycle. 6 (13): 1553–7. PMID 17611415.
- Mullor JL, Sánchez P, Altaba AR (2003). "Pathways and consequences: Hedgehog signaling in human disease". Trends Cell Biol. 12 (12): 562–9. doi:10.1016/S0962-8924(02)02405-4. PMID 12495844.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Nanni L, Ming JE, Du Y; et al. (2001). "SHH mutation is associated with solitary median maxillary central incisor: a study of 13 patients and review of the literature". Am. J. Med. Genet. 102 (1): 1–10. doi:10.1002/1096-8628(20010722)102:1<1::AID-AJMG1336>3.0.CO;2-U. PMID 11471164.
{{cite journal}}: Explicit use of et al. in:|author=(help)CS1 maint: multiple names: authors list (link) - Williams JA (2006). "Hedgehog and spinal cord injury". Expert Opin. Ther. Targets. 9 (6): 1137–45. doi:10.1517/14728222.9.6.1137. PMID 16300466.
External links
- An introductory article on shh at Davidson College
- SHH at The GDB Human Genome Database
- Rediscovering biology: Unit 7, Genetics of development. Expert interview transcripts, interview with John Incardona, PhD. explanation of the discovery and naming of the sonic hedgehog gene
- ‘Sonic Hedgehog’ sounded funny, at first. New York Times, November 12, 2006.
PDB gallery | |
|---|---|
|
