Microscopic image of the epidermis, which constitutes the outer layer of skin, shown here by the white bar
Microscopic image showing the layers of the epidermis. The stratum corneum appears more compact in this image than above because of different sample preparation.
The word epidermis is derived through Latin from Ancient Greek epidermis, itself from Ancient Greek epi 'over, upon' and from Ancient Greek derma 'skin'. Something related to or part of the epidermis is termed epidermal.
Epidermal cells are tightly interconnected to serve as a tight barrier against the exterior environment. The junctions between the epidermal cells are of the adherens junction type, formed by transmembrane proteins called cadherins. Inside the cell, the cadherins are linked to actin filaments. In immunofluorescence microscopy, the actin filament network appears as a thick border surrounding the cells,[9] although the actin filaments are actually located inside the cell and run parallel to the cell membrane. Because of the proximity of the neighboring cells and tightness of the junctions, the actin immunofluorescence appears as a border between cells.[9]
Confocal image of the stratum corneumComposed of 10 to 30 layers of polyhedral, anucleated corneocytes (final step of keratinocyte differentiation), with the palms and soles having the most layers. Corneocytes contain a protein envelope (cornified envelope proteins) underneath the plasma membrane, are filled with water-retaining keratin proteins, attached together through corneodesmosomes and surrounded in the extracellular space by stacked layers of lipids.[11] Most of the barrier functions of the epidermis localize to this layer.[12]
clear/translucent layer (stratum lucidum, only in palms and soles)
This narrow layer is found only on the palms and soles. The epidermis of these two areas is known as "thick skin" because with this extra layer, the skin has 5 epidermal layers instead of 4.
Confocal image of the stratum granulosumKeratinocytes lose their nuclei and their cytoplasm appears granular. Lipids, contained into those keratinocytes within lamellar bodies, are released into the extracellular space through exocytosis to form a lipid barrier that prevents water loss from the body as well as entry of foreign substances. Those polar lipids are then converted into non-polar lipids and arranged parallel to the cell surface. For example glycosphingolipids become ceramides and phospholipids become free fatty acids.[11]
Confocal image of the stratum spinosum already showing some clusters of basal cellsKeratinocytes become connected through desmosomes and produce lamellar bodies, from within the Golgi, enriched in polar lipids, glycosphingolipids, free sterols, phospholipids and catabolic enzymes.[3] Langerhans cells, immunologically active cells, are located in the middle of this layer.[11]
Confocal image of the stratum basale already showing some papillaeComposed mainly of proliferating and non-proliferating keratinocytes, attached to the basement membrane by hemidesmosomes. Melanocytes are present, connected to numerous keratinocytes in this and other strata through dendrites. Merkel cells are also found in the stratum basale with large numbers in touch-sensitive sites such as the fingertips and lips. They are closely associated with cutaneous nerves and seem to be involved in light touch sensation.[11]
As a stratified squamous epithelium, the epidermis is maintained by cell division within the stratum basale. Differentiating cells delaminate from the basement membrane and are displaced outward through the epidermal layers, undergoing multiple stages of differentiation until, in the stratum corneum, losing their nucleus and fusing to squamous sheets, which are eventually shed from the surface (desquamation). Differentiated keratinocytes secrete keratin proteins, which contribute to the formation of an extracellular matrix that is an integral part of the skin barrier function. In normal skin, the rate of keratinocyte production equals the rate of loss,[4] taking about two weeks for a cell to journey from the stratum basale to the top of the stratum granulosum, and an additional four weeks to cross the stratum corneum.[2] The entire epidermis is replaced by new cell growth over a period of about 48 days.[13]
Keratinocyte differentiation throughout the epidermis is in part mediated by a calcium gradient, increasing from the stratum basale until the outer stratum granulosum, where it reaches its maximum, and decreasing in the stratum corneum. Calcium concentration in the stratum corneum is very low in part because those relatively dry cells are not able to dissolve the ions. This calcium gradient parallels keratinocyte differentiation and as such is considered a key regulator in the formation of the epidermal layers.[3]
Elevation of extracellular calcium concentrations induces an increase in intracellular free calcium concentrations.[14] Part of that intracellular increase comes from calcium released from intracellular stores[15] and another part comes from transmembrane calcium influx,[16] through both calcium-sensitive chloride channels[17] and voltage-independent cation channels permeable to calcium.[18] Moreover, it has been suggested that an extracellular calcium-sensing receptor (CaSR) also contributes to the rise in intracellular calcium concentration.[19]
The cells in the stratum granulosum do not divide, but instead form skin cells called keratinocytes from the granules of keratin. These skin cells finally become the cornified layer (stratum corneum), the outermost epidermal layer, where the cells become flattened sacks with their nuclei located at one end of the cell. After birth these outermost cells are replaced by new cells from the stratum granulosum and throughout life they are shed at a rate of 30 - 90 milligrams of skin flakes every hour, or 0.720 - 2.16 grams per day.[21]
The epidermis serves as a barrier to protect the body against microbial pathogens, oxidant stress (UV light), and chemical compounds, and provides mechanical resistance to minor injury. Most of this barrier role is played by the stratum corneum.[12]
Characteristics
Physical barrier: Epidermal keratinocytes are tightly linked by cell–cell junctions associated to cytoskeletal proteins, giving the epidermis its mechanical strength.[3]
Chemical barrier: Highly organized lipids, acids, hydrolytic enzymes, and antimicrobial peptides[3] inhibit passage of external chemicals and pathogens into the body.
Immunologically active barrier: The humoral and cellular constituents of the immune system[3] found in the epidermis actively combat infection.
Water content of the stratum corneum drops towards the surface, creating hostile conditions for pathogenic microorganism growth.[12]
An acidic pH (around 5.0) and low amounts of water make the epidermis hostile to many microorganic pathogens.[12]
Non-pathogenic microorganisms on the surface of the epidermis help defend against pathogens by competing for food, limiting its availability, and producing chemical secretions that inhibit the growth of pathogenic microbiota.[12]
The ability of the skin to hold water is primarily due to the stratum corneum and is critical for maintaining healthy skin.[24] Skin hydration is quantified using corneometry.[25] Lipids arranged through a gradient and in an organized manner between the cells of the stratum corneum form a barrier to transepidermal water loss.[26][27]
The amount and distribution of melaninpigment in the epidermis is the main reason for variation in skin color in Homo sapiens. Melanin is found in the small melanosomes, particles formed in melanocytes from where they are transferred to the surrounding keratinocytes. The size, number, and arrangement of the melanosomes vary between racial groups, but while the number of melanocytes can vary between different body regions, their numbers remain the same in individual body regions in all human beings. In white and Asian skin the melanosomes are packed in "aggregates", but in black skin they are larger and distributed more evenly. The number of melanosomes in the keratinocytes increases with UV radiation exposure, while their distribution remain largely unaffected.[28]
The skin contains specialized epidermal touch receptor cells called Merkel cells. Historically, the role of Merkel cells in sensing touch has been thought to be indirect, due their close association with nerve endings. However, recent work in mice and other model organisms demonstrates that Merkel cells intrinsically transform touch into electrical signals that are transmitted to the nervous system.[29]
Laboratory culture of keratinocytes to form a 3D structure (artificial skin) recapitulating most of the properties of the epidermis is routinely used as a tool for drug development and testing.
Focal epithelial hyperplasia (Heck's disease) is an asymptomatic, benign neoplastic condition characterized by multiple white to pinkish papules that occur diffusely in the oral cavity.[32][6]: 411
Pseudoepitheliomatous hyperplasia (PEH) is a benign condition characterized by hyperplasia of the epidermis and epithelium of skin appendages,[33] with irregular squamous strands extending down into the dermis,[34] and closely simulating squamous cell carcinoma (SCC).[33]
Pseudoepitheliomatous hyperplasia (PEH), low magnification, with acanthotic squamous epithelium with irregular thick finger-like downgrowths into the underlying dermis.
PEH, high magnification, with reactive-appearing squamous downgrowths with no significant cytologic atypia.
^Hennings H, Kruszewski FH, Yuspa SH, Tucker RW (April 1989). "Intracellular calcium alterations in response to increased external calcium in normal and neoplastic keratinocytes". Carcinogenesis. 10 (4): 777–780. doi:10.1093/carcin/10.4.777. PMID2702726.
^Pillai S, Bikle DD (January 1991). "Role of intracellular-free calcium in the cornified envelope formation of keratinocytes: differences in the mode of action of extracellular calcium and 1,25 dihydroxyvitamin D3". Journal of Cellular Physiology. 146 (1): 94–100. doi:10.1002/jcp.1041460113. PMID1990023. S2CID21264605.
^Reiss M, Lipsey LR, Zhou ZL (May 1991). "Extracellular calcium-dependent regulation of transmembrane calcium fluxes in murine keratinocytes". Journal of Cellular Physiology. 147 (2): 281–291. doi:10.1002/jcp.1041470213. PMID1645742. S2CID25858560.
^Mauro TM, Pappone PA, Isseroff RR (April 1990). "Extracellular calcium affects the membrane currents of cultured human keratinocytes". Journal of Cellular Physiology. 143 (1): 13–20. doi:10.1002/jcp.1041430103. PMID1690740. S2CID8072916.
^Mauro TM, Isseroff RR, Lasarow R, Pappone PA (March 1993). "Ion channels are linked to differentiation in keratinocytes". The Journal of Membrane Biology. 132 (3): 201–209. doi:10.1007/BF00235738. PMID7684087. S2CID13063458.
^Blichmann CW, Serup J (1988). "Assessment of skin moisture. Measurement of electrical conductance, capacitance and transepidermal water loss". Acta Dermato-venereologica. 68 (4): 284–90. doi:10.2340/0001555568284290 (inactive 31 January 2024). PMID2459872.{{cite journal}}: CS1 maint: DOI inactive as of January 2024 (link)
^Downing DT, Stewart ME, Wertz PW, Colton SW, Abraham W, Strauss JS (March 1987). "Skin lipids: an update". The Journal of Investigative Dermatology. 88 (3 Suppl): 2s–6s. doi:10.1111/1523-1747.ep12468850 (inactive 2024-02-01). PMID2950180.{{cite journal}}: CS1 maint: DOI inactive as of February 2024 (link)
^Bonté F, Saunois A, Pinguet P, Meybeck A (January 1997). "Existence of a lipid gradient in the upper stratum corneum and its possible biological significance". Archives of Dermatological Research. 289 (2): 78–82. doi:10.1007/s004030050158. PMID9049040. S2CID10787600.
^Kumar V, Fausto N, Abbas A (2004). Robbins & Cotran Pathologic Basis of Disease (7th ed.). Saunders. p. 1230. ISBN0-7216-0187-1.
^Stone MS, Ray TL (September 1995). "Acanthosis". DermPathTutor. Department of Dermatology, University of Iowa. Archived from the original on 29 May 2012. Retrieved 17 May 2012.







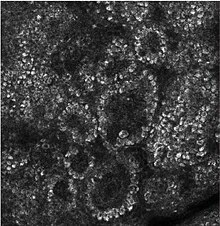






 Pseudoepitheliomatous hyperplasia (PEH), low magnification, with acanthotic squamous epithelium with irregular thick finger-like downgrowths into the underlying dermis.
Pseudoepitheliomatous hyperplasia (PEH), low magnification, with acanthotic squamous epithelium with irregular thick finger-like downgrowths into the underlying dermis.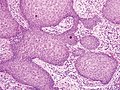 PEH, high magnification, with reactive-appearing squamous downgrowths with no significant cytologic atypia.
PEH, high magnification, with reactive-appearing squamous downgrowths with no significant cytologic atypia.



 Epidermis and dermis of human skin
Epidermis and dermis of human skin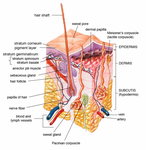 Cross-section of all skin layers
Cross-section of all skin layers Illustration of epidermal layers
Illustration of epidermal layers Optical coherence tomography of fingertip
Optical coherence tomography of fingertip


