
Thalamus: Difference between revisions
m Reverted edits by 24.107.37.17 (talk) editing tests or vandalism (HG) |
Tag: Removal of interwiki link; Wikidata is live |
||
| Line 125: | Line 125: | ||
==External links== |
==External links== |
||
{{Commons category| |
{{Commons category|Thalaeghslkdiughleriuhyger6eopiopjdupioty |
||
{{wiktionarypar|thalamus}} |
|||
* http://www.scholarpedia.org/article/Thalamus |
|||
* {{BrainMaps|thalamus}} |
|||
{{Diencephalon}} |
|||
{{Neural tracts}} |
|||
[[Category:Diencephalon]] |
|||
[[ar:مهاد]] |
|||
[[bg:Таламус]] |
|||
[[ca:Tàlem]] |
|||
[[cs:Thalamus]] |
|||
[[da:Thalamus]] |
|||
[[de:Thalamus]] |
|||
[[et:Taalamus]] |
|||
[[es:Tálamo (SNC)]] |
|||
[[eu:Talamo]] |
|||
[[fa:تالاموس]] |
|||
[[fr:Thalamus]] |
|||
[[gl:Tálamo]] |
|||
[[ko:시상]] |
|||
[[it:Talamo]] |
|||
[[he:תלמוס]] |
|||
[[kk:Көру төмпегі, таламус]] |
|||
[[lt:Gumburas]] |
|||
[[hu:Talamusz]] |
|||
[[nl:Thalamus]] |
|||
[[ja:視床]] |
|||
[[no:Talamus]] |
|||
[[pl:Wzgórze (anatomia)]] |
|||
[[pt:Tálamo (anatomia)]] |
|||
[[ro:Talamus]] |
|||
[[ru:Таламус]] |
|||
[[sr:Таламус]] |
|||
[[fi:Talamus]] |
|||
[[sv:Talamus]] |
|||
[[th:ทาลามัส]] |
|||
[[tr:Talamus]] |
|||
[[uk:Таламус]] |
|||
[[ur:مہاد]] |
|||
[[zh:丘脑]] |
|||
Revision as of 15:44, 12 October 2011
| Thalamus | |
|---|---|
 MRI cross-section of human brain, with thalamus marked. | |
 Scheme showing the parts of the thalamus | |
| Details | |
| Part of | Diencephalon |
| Parts | See List of thalamic nuclei |
| Identifiers | |
| Latin | thalamus dorsalis |
| MeSH | D013788 |
| NeuroNames | 300 |
| NeuroLex ID | birnlex_954 |
| TA98 | A14.1.08.101 A14.1.08.601 |
| TA2 | 5678 |
| TE | E5.14.3.4.2.1.8 |
| FMA | 62007 |
| Anatomical terms of neuroanatomy | |
The thalamus (from Greek θάλαμος = room, chamber) is a midline paired symmetrical structure within the brains of vertebrates, including humans. It is situated between the cerebral cortex and midbrain, both in terms of location and neurological connections. Its function includes relaying sensation, spatial sense, and motor signals to the cerebral cortex, along with the regulation of consciousness, sleep, and alertness. The thalamus surrounds the third ventricle. It is the main product of the embryonic diencephalon.
Location and topography

The thalamus is the largest structure in the diencephalon, the part of the brain situated between the midbrain (mesencephalon) and forebrain (telencephalon). Anatomically, the thalamus is perched on top of the brainstem, near the center of the brain, in a position to send nerve fibers out to the cerebral cortex in all directions. The diencephalon includes also the dorsally located epithalamus (essentially the habenula and annexes) and the perithalamus (prethalamus formerly described as ventral thalamus) containing the zona incerta and the "reticulate nucleus" (not the reticular, term of confusion). Due to their different ontogenetic origins, the epithalamus and the perithalamus are formally distinguished from the thalamus proper.
In humans, the two halves of the thalamus are prominent bulb-shaped masses, about 5.7 cm in length, located obliquely (about 30°) and symmetrically on each side of the third ventricle.
Anatomy

The thalamus comprises a system of lamellae (made up of myelinated fibers) separating different thalamic subparts. Other areas are defined by distinct clusters of neurons, such as the periventricular gray, the intralaminar elements, the "nucleus limitans", and others.[1] These latter structures, different in structure from the major part of the thalamus, have been grouped together into the allothalamus as opposed to the isothalamus.[2] This distinction simplifies the global description of the thalamus.
See also List of thalamic nuclei.
Arterial supply
The thalamus derives its blood supply from four arteries including the polar artery(posterior communicating artery), paramedian thalamic-subthalamic arteries, inferolateral (thalamogeniculate) arteries, and posterior (medial and lateral) choroidal arteries.[3] These are all derived from the vertebrobasilar arterial system except the polar artery.
Function
The thalamus has multiple functions. It is generally believed to act as a relay between a variety of subcortical areas and the cerebral cortex. In particular, every sensory system (with the exception of the olfactory system) includes a thalamic nucleus that receives sensory signals and sends them to the associated primary cortical area. For the visual system, for example, inputs from the retina are sent to the lateral geniculate nucleus of the thalamus, which in turn projects to the primary visual cortex (area V1) in the occipital lobe. The thalamus is believed to both process sensory information as well as relay it—each of the primary sensory relay areas receives strong "back projections" from the cerebral cortex. Similarly the medial geniculate nucleus acts as a key auditory relay between the inferior colliculus of the midbrain and the primary auditory cortex, and the ventral posterior nucleus is a key somatosensory relay, which sends touch and proprioceptive information to the primary somatosensory cortex.
The thalamus also plays an important role in regulating states of sleep and wakefulness.[4] Thalamic nuclei have strong reciprocal connections with the cerebral cortex, forming thalamo-cortico-thalamic circuits that are believed to be involved with consciousness. The thalamus plays a major role in regulating arousal, the level of awareness, and activity. Damage to the thalamus can lead to permanent coma.
Many different functions are linked to various regions of the thalamus. This is the case for many of the sensory systems (except for the olfactory system), such as the auditory, somatic, visceral, gustatory and visual systems where localized lesions provoke specific sensory deficits. A major role of the thalamus is devoted to "motor" systems. This has been and continues to be a subject of interest for investigators. VIm, the relay of cerebellar afferences, is the target of stereotactians particularly for the improvement of tremor. The role of the thalamus in the more anterior pallidal and nigral territories in the basal ganglia system disturbances is recognized but still poorly understood. The contribution of the thalamus to vestibular or to tectal functions is almost ignored. The thalamus has been thought of as a "relay" that simply forwards signals to the cerebral cortex. Newer research suggests that thalamic function is more selective.[5]
Pathology
A cerebrovascular accident (stroke) can lead to the thalamic syndrome,[6] which involves a one-sided burning or aching sensation often accompanied by mood swings. Bilateral ischemia of the area supplied by the paramedian artery can cause serious problems including akinetic mutism, and be accompanied by oculomotor problems. A related concept is thalamocortical dysrhythmia.
Korsakoff's syndrome stems from damage to the mammillary body, the mammillothalamic fasciculus or the thalamus.
Fatal familial insomnia is a hereditary prion disease in which degeneration of the thalamus occurs, causing the patient to gradually lose his ability to sleep and progressing to a state of total insomnia, which invariably leads to death.
Development
The thalamic complex is composed of the perithalamus (or prethalamus, previously also known as ventral thalamus), the mid-diencephalic organiser (which forms later the zona limitans intrathalamica (ZLI) ) and the thalamus (dorsal thalamus).[7][8] The development of the thalamus can be subdivide into three steps[9]
Early brain development
After neurulation the anlage of the prethalamus and the thalamus is induced within the neural tube. Data from different vertebrate model organisms support a model in which the interaction between two transcription factors, Fez and Otx, are of decisive importance. Fez is expressed in the prethalamus, and functional experiments show that Fez is required for prethalamus formation.[10][11] Posteriorly, Otx1 and Otx2 abut the expression domain of Fez and are required for proper development of the thalamus.[12][13]
The formation of the mid-diencephalic organiser (MDO)
At the interface between the expression domains of Fez and Otx, the mid-diencephalic organizer (MDO, also called the ZLI organiser) is induced within the thalamic anlage. The MDO is the central signalling organizer in the thalamus. A lack of the organizer leads to the absence of the thalamus. The MDO matures from ventral to dorsal during development. Members of the SHH family and of the Wnt family are the main principal signals emitted by the MDO.
Besides its importance as signalling center, the organizer matures into the morphological structure of the zona limitans intrathalamica (ZLI).
Maturation and parcellation of the thalamus
After its induction, the MDO starts to orchestrate the development of the thalamic anlage by release of signalling molecules such as Shh.[14] In mice, the function of signaling at the MDO has not been addressed directly due to a complete absence of the diencephalon in Shh mutants.[15]
Studies in chicks have shown that Shh is both necessary and sufficient for thalamic gene induction.[16] In zebrafish, it was shown that the expression of two Shh genes, shh-a and shh-b (formerly described as twhh) mark the MDO territory, and that Shh signaling is sufficient for the molecular differentiation of both the prethalamus and the thalamus but is not required for their maintenance and Shh signaling from the MDO/alar plate is sufficient for the maturation of prethalamic and thalamic territory while ventral Shh signals are dispensable.[17] The exposure to Shh leads to differentiation of thalamic neurons. SHH signaling from the MDO induces a posterior-to-anterior wave of expression the proneural gene Neurogenin1 in the major (caudal) part of the thalamus, and Ascl1 (formerly Mash1) in the remaining narrow stripe of rostral thalamic cells immediately adjacent to the MDO, and in the prethalamus[18][19]
This zonation of proneural gene expression leads to the differentiation of glutamatergic relay neurons from the Neurogenin1+ precursors and of GABAergic inhibitory neurons from the Ascl1+ precursors. In fish, selection of these alternative neurotransmitter fates is controlled by the dynamic expression of Her6 the homolog of HES1. Expression of this hairy-like bHLH transcription factor, which represses Neurogenin but is required for Ascl1, is progressively lost from the caudal thalamus but maintained in the prethalamus and in the stripe of rostral thalamic cells. In addition, studies on chick and mice have shown that blocking the Shh pathway leads to absence of the rostral thalamus and substantial decrease of the caudal thalamus. The rostral thalamus will give rise to the reticular nucleus mainly whereby the caudal thalamus will form the relay thalamus and will be further subdivided in the thalamic nuclei.
Further reading: Building a bridal chamber: development of the thalamus. from Steffen Scholpp and Andrew Lumsden in Trends of Neuroscience 2010[20]
In humans, a common genetic variation in the promotor region of the serotonin transporter (the SERT-long and -short allele: 5-HTTLPR) has been shown to affect the development of several regions of the thalamus in adults. People who inherit two short alleles (SERT-ss) have more neurons and a larger volume in the pulvinar and possibly the limbic regions of the thalamus. Enlargement of the thalamus provides an anatomical basis for why people who inherit two SERT-ss alleles are more vulnerable to major depression, posttraumatic stress disorder, and suicide.[21]
See also
- Primate basal ganglia system
- List of regions in the human brain
- Thalamus (non primate)
- List of thalamic nuclei
- Thalamic stimulator
- Thalamotomy
References
- ^ Jones Edward G.(2007) "The Thalamus" Cambridge Uni. Press
- ^ Percheron, G. (2003) "Thalamus". In Paxinos, G. and May, J.(eds). The human nervous system. 2d Ed. Elsevier. Amsterdam. pp.592-675
- ^ Percheron, G. (1982) The arterial supply of the thalamus. In Schaltenbrand and Walker, A.E.(eds) Stereotaxy of the human brain. Thieme . Stuttgart. pp.218-232
- ^ Steriade, M. and Llinas, R. (1988) "The functional states of the thalamus and the associated neuronal interplay". Physiological Reviews 68: 699-742 PMID 2839857
- ^ Your Brain Boots Up Like a Computer | LiveScience
- ^ Dejerine, J. and Roussy, G. (1906) Le syndrome thalamique. Rev. Neurol. 14: 521-532
- ^ Kuhlenbeck, H. (1937). The ontogenetic development of diencephalic centres in the bird's brain (chick) and comparison with the reptilian and mammalian diencephalon. J. Comp. Neurol. 66
- ^ Shimamura, K., Hartigan, D. J., Martinez, S., Puelles, L. and Rubenstein, J. L. (1995). Longitudinal organization of the anterior neural plate and neural tube. Development 121,3923 -3933.
- ^ Scholpp S, Lumsden A Building a bridal chamber: development of the thalamus. Trends Neurosci. 2010 Aug;33(8):373-380. Epub 2010 Jun 11. [1]
- ^ 8 Hirata, T., et al. (2006) Zinc-finger genes Fez and Fez-like function in the establishment of diencephalon subdivisions. Development 133, 3993-4004 [2]
- ^ Jeong JY, Einhorn Z, Mathur P, Chen L, Lee S, Kawakami K, Guo S. Patterning the zebrafish diencephalon by the conserved zinc-finger protein Fezl. Development. 2007 Jan;134(1):127-36. [3]
- ^ Acampora D, Avantaggiato V, Tuorto F, Simeone A. Genetic control of brain morphogenesis through Otx gene dosage requirement. Development. 1997 Sep;124(18):3639-50. [4]
- ^ Scholpp S, Foucher I, Staudt N, Peukert D, Lumsden A, Houart C. Otx1l, Otx2 and Irx1b establish and position the ZLI in the diencephalon. Development. 2007 Sep;134(17):3167-76. [5]
- ^ Puelles, L. and Rubenstein, J. L. (2003). Forebrain gene expression domains and the evolving prosomeric model. Trends Neurosci. 26,469 -476.
- ^ Ishibashi, M. and McMahon, A. P. (2002). A sonic hedgehog-dependent signalling relay regulates growth of diencephalic and mesencephalic primordia in the early mouse embryo. Development 129,4807 -4819.
- ^ Kiecker, C. and Lumsden, A. (2004). Hedgehog signalling from the ZLI regulates diencephalic regional identity. Nat. Neurosci. 7,1242 -1249.
- ^ Scholpp S, Wolf O, Brand M, Lumsden A. Hedgehog signalling from the zona limitans intrathalamica orchestrates patterning of the zebrafish diencephalon'. Development. 2006 Mar;133(5):855-64 [6]
- ^ Scholpp S, Delogu A, Gilthorpe J, Peukert D, Schindler S, Lumsden A. Her6 regulates the neurogenetic gradient and neuronal identity in the thalamus. Proc Natl Acad Sci U S A. 2009 Nov 24;106(47):19895-900 [7]
- ^ Vue TY, Bluske K, Alishahi A, Yang LL, Koyano-Nakagawa N, Novitch B, Nakagawa Y. Sonic hedgehog signaling controls thalamic progenitor identity and nuclei specification in mice. J Neurosci. 2009 Apr 8;29(14):4484-97 [8]
- ^ Scholpp S, Lumsden A Building a bridal chamber: development of the thalamus. Trends Neurosci. 2010 Aug;33(8):373-380. Epub 2010 Jun 11. [9]
- ^ Young KA, Holcomb LA, Yazdani U, Bonkale W, Hicks PB and German DC (2007). "5HTTLPR polymorphism and enlargement of the pulvinar: Unlocking the backdoor to the limbic system". Biol Psychiatry. 61 (6): 813–8. doi:10.1016/j.biopsych.2006.08.047. PMID 17083920.
{{cite journal}}: CS1 maint: multiple names: authors list (link)
Additional images
-
 Schematic representation of the chief ganglionic categories (I to V).
Schematic representation of the chief ganglionic categories (I to V). -
 Deep dissection of brain-stem. Lateral view.
Deep dissection of brain-stem. Lateral view. -
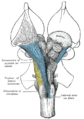 Deep dissection of brain-stem. Ventral view.
Deep dissection of brain-stem. Ventral view. -
 Mesal aspect of a brain sectioned in the median sagittal plane.
Mesal aspect of a brain sectioned in the median sagittal plane. -
 Dissection showing the ventricles of the brain.
Dissection showing the ventricles of the brain. -
 Coronal section of brain immediately in front of pons.
Coronal section of brain immediately in front of pons. -
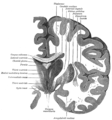 Coronal section of brain through intermediate mass of third ventricle.
Coronal section of brain through intermediate mass of third ventricle. -
 Coronal section of lateral and third ventricles.
Coronal section of lateral and third ventricles. -
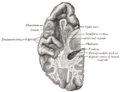 Section of brain showing upper surface of temporal lobe.
Section of brain showing upper surface of temporal lobe. -
 Horizontal section of right cerebral hemisphere.
Horizontal section of right cerebral hemisphere. -
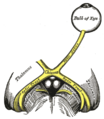 The left optic nerve and the optic tracts.
The left optic nerve and the optic tracts. -
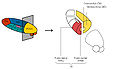 Parts of the thalamus
Parts of the thalamus -
 Projections of the thalamus
Projections of the thalamus -
 Parts of the thalamus
Parts of the thalamus -
 Thalamus
Thalamus -
 Human brain frontal (coronal) section
Human brain frontal (coronal) section -

-

-
 Nuclei of the thalamus
Nuclei of the thalamus















_section_description.JPG)



External links
{{Commons category|Thalaeghslkdiughleriuhyger6eopiopjdupioty