
Ochnaceae: Difference between revisions
I added to the taxonomy section. |
I mentioned 2 classifications from 2014. |
||
| Line 141: | Line 141: | ||
Medusagynoideae consists of a single species, ''Medusagyne oppositifolia''. It is [[endemic]] to the [[island]] of [[Mahé]] in the [[Seychelles]].<ref name=dickison1990>William C. Dickison. 1990. "The morphology and relationships of ''Medusagyne'' (Medusagynaceae)". ''Plant Systematics and Evolution'' '''171'''(1-4):27-55.</ref> |
Medusagynoideae consists of a single species, ''Medusagyne oppositifolia''. It is [[endemic]] to the [[island]] of [[Mahé]] in the [[Seychelles]].<ref name=dickison1990>William C. Dickison. 1990. "The morphology and relationships of ''Medusagyne'' (Medusagynaceae)". ''Plant Systematics and Evolution'' '''171'''(1-4):27-55.</ref> |
||
Quiinoideae comprises about 48 species in four genera: ''Froesia'', ''Quiina'', ''Touroulia'', and ''Lacunaria''. It is restricted to [[tropical]] [[Americas|America]].<ref name=schneiderinpress>Julio V. Schneider and Georg Zizka. in press. "Quiinaceae". ''Flora Neotropica'' monograph ??.</ref> |
Quiinoideae comprises about 48 species in four genera: ''Froesia'', ''Quiina'', ''Touroulia'', and ''Lacunaria''. It is restricted to [[tropical]] [[Americas|America]].<ref name=schneiderinpress>Julio V. Schneider and Georg Zizka. in press. "Quiinaceae". ''Flora Neotropica'' monograph ??.</ref> ''Froesia'' is distinct from the other three genera. Its flowers are always bisexual, and the fruit consists of three structures that resemble [[Follicle (fruit)|follicles]], except that they are not quite entirely separate from each other. |
||
The subfamily Ochnoideae was revised in 2014 as Ochnaceae [[sensu stricto]].<ref name=amaral2014/> In that treatment, 27 genera were described. An additional genus, ''Neckia'', was resurrected in that same year, based on the results of a molecular phylogenetic study.<ref name=schneider2014/> Amaral and Bittrich (2014) divided Ochnoideae into three tribes: Luxemburgieae, Sauvagesieae, and Ochneae. The genus ''Testulea'' was included in the tribe Sauvagesieae. No subtribes were recognized. |
|||
In 2014, a second reclassification of Ochnoideae was published, based on [[cladistic]] analysis of [[DNA sequence]]s. In that paper, Schneider [[et alii]] divided Ochnoideae into four tribes: Testuleeae, Luxemburgieae, Sauvagesieae, and Ochneae. Their circumscription of the tribes was the same as that of Amaral and Bittrich (2014), except that ''Testulea'' was removed from Sauvagesieae into its own tribe, Testuleeae. Inclusion of ''Testulea'' in Sauvagesieae renders that tribe [[paraphyletic]] over Luxemburgieae. |
|||
In 1902, [[Philippe van Tieghem]] recognized the following eight families in what is now Ochnaceae [[sensu lato]]: |
In 1902, [[Philippe van Tieghem]] recognized the following eight families in what is now Ochnaceae [[sensu lato]]: |
||
Revision as of 04:58, 1 May 2015
This article currently links to a large number of disambiguation pages (or back to itself). (April 2015) |
| Ochnaceae | |
|---|---|

| |
| Sauvagesia erecta from southern Brazil | |
| Scientific classification | |
| Kingdom: | |
| (unranked): | |
| (unranked): | |
| (unranked): | |
| Order: | |
| Family: | Ochnaceae |
| Genera | |
|
See Subdivisions | |
This article or section is in a state of significant expansion or restructuring. You are welcome to assist in its construction by editing it as well. If this article or section has not been edited in several days, please remove this template. If you are the editor who added this template and you are actively editing, please be sure to replace this template with {{in use}} during the active editing session. Click on the link for template parameters to use.
This article was last edited by 128.171.106.250 (talk | contribs) 9 years ago. (Update timer) |
Ochnaceae is a family of flowering plants in the order Malpighiales.[2] In the APG III system of classification of flowering plants, Ochnaceae is defined broadly, to include about 550 species,[3] and encompasses what some taxonomists have treated as the separate families Medusagynaceae and Quiinaceae.[1] In a phylogenetic study that was published in 2014, Ochnaceae was recognized in the broad sense,[4] but two works published after APG III have accepted the small families Medusagynaceae and Quiinaceae.[3][5]
In this article, "Ochnaceae" will refer to the larger circumscription of the family, which is otherwise known as Ochnaceae sensu lato or as the ochnoids.[6]
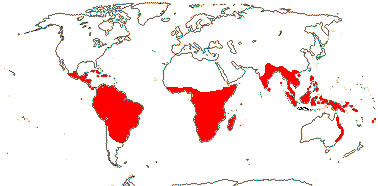
Ochnaceae, defined broadly or narrowly, is pantropical in distribution, with a few species cultivated outside of this range. Ochnaceae is most diverse in the neotropics, with a second center of diversity in tropical Africa.[4] It consists mostly of shrubs and small trees, and a few herbaceous species in Sauvagesia. Many are treelets, with a single, erect trunk, but low in height. The Ochnaceae are notable for their unusual leaves. These are usually shiny, with closely spaced, parallel veins, toothed margins, and conspicuous stipules. Most of the species are buzz pollinated.[7] In eight of the genera in tribe Sauvagesieae, the flower changes form after opening, by continued growth of tissue within the flower.[4]
A few species of Ochna are cultivated as ornamentals.[8] Ochna thomasiana is probably the most commonly planted, but it is often misidentified in the horticultural literature.[9]
The leaves of Cespedesia are sometimes to 1 m (3.3 ft) in length and are used for roofing.[10] An herbal tea is made from the pantropical weed Sauvagesia erecta.
In its evolution, Ochnaceae has been unusual, in "reverting" to character states that are regarded as ancestral or primitive. For example, an actinomorphic floral symmetry has appeared twice in the subfamily Ochnoideae. Also, two clades of Ochnaceae, one in Ochnoideae and another in Quiinoideae have a derived condition very close to apocarpy. The complete separation of the carpels (apocarpy) is thought to be the ancestral state for angiosperms.[11]
Fossils attributed to Ochnaceae are known from the early Eocene of Mississippi.[12] The age of the family is very roughly estimated at 100 million years.[13]
A great many genus names have been published in Ochnaceae.[14] In a taxonomic revision of Ochnaceae, as three families, in 2014, only 32 of these genera were accepted; one in Medusagynaceae, four in Quiinaceae, and 27 in Ochnaceae s.s..[3] In that same year, a 33rd genus, Neckia, was reestablished in order to preserve the monophyly of another genus, Sauvagesia.[4]
The largest genera in Ochnaceae are: Ouratea (200 species), Ochna (85), Campylospermum (65), Sauvagesia (39), and Quiina (34).[3] None of the larger genera has been the subject of a phylogenetic analysis of DNA sequences of selected genes. In one study of the subfamily Quiinoideae, based on the trn L-F intergenic spacer, only nine species were sampled from this subfamily.[15]
Genera
The following list of 33 genera consists of Neckia, which was resurrected in 2014,[4] plus the 32 genera that were described in the most recent revision of Ochnaceae.[3][16][17] The classification is from Schneider et alii (2014).[4]
|
|
Description
The following description is excerpted from the descriptions of Medusagynaceae, Quiinaceae, and Ochnaceae s.s. in The Families and Genera of Vascular Plants,[17][16][3] with some information from other sources, as indicated.
Mostly shrubs and small trees, plus a few trees of moderate size, and in Sauvagesia, a few herbs.
Leaves simple, except pinnately compound in Krukoviella,[2] and in Quiinoideae, often pinnately lobed or compound on juvenile plants. Pinnate leaves are typical of Rhytidanthera.[4] The leaves are often coriaceous and conspicuously serrate. Stipules present, except Medusagyne.
Venation often scalariform (ladder-like) in appearance, with parallel and closely spaced secondary and tertiary veins. Petioles absent or short, sometimes resembling a pulvinus.[2]
Unisexual flowers are common in Medusagyne and in Quiinoideae (except Froesia), but restricted to a clade of three genera in Ochnoideae. Unisexual flowers are found in Schuurmansia, Schuurmansiella, and Euthemis.[4] The flowers are always unisexual in Schuurmansiella.[3]
In polygamous species, flowers have been assessed as bisexual on the basis of morphology only.[15] Pollen produced by apparently hermaphroditic flowers has, in a few cases, been found to be inaperturate, rendering the flower functionally female.
Sepals 3 to 5, often unequal, sometimes accrescent.
Petals 4 or 5,, or rarely 3, 6, 7, or 8,, often contort, free or fuzed at base only, sometimes reflexed over the sepals.
Fertile stamens 5 to 10 or numerous, rarely one. Filaments sometimes persistent, sometimes narrowed near the anthers.
Anthers basifixed or slightly dorsifixed, usually dehiscing by one or two apical or subapical pores, sometimes latrorsely by longitudinal slits. In Medusagyne and Quiinoideae, an enlarged septum separates the thecae.
Staminodes often present, free or connate, sometimes petaloid, sometimes enveloping the fertile stamens.
Nectar not produced. Flowers usually buzz-pollinated.
Ovary superior, longitudinally ribbed in Medusagyne and Quiinoideae; unribbed in Ochnoideae. Carpels completely fused or nearly separate; 2-15, or up to 25 in Medusagyne. Style apical or gynobasic.
Fruit sometimes winged; rarely a nut or drupe, often berry-like; usually a septicidal capsule, or else the ovary separating to form blackish drupelets on a usually reddish, accrescent receptacle.
Seeds albuminous or exalbuminous, winged or not. The seed coat often includes a layer of cristarque cells. These are sclereids, each containing calcium oxalate crystals in the form of a druse.
Classification
Until near the end of the 20th century, Ochnaceae was regarded as a rather odd family, difficult to place with a high degree of certainty. Even into the 21st century, some authors treated the genus Strasburgeria as the closest relative of Ochnaceae, and some even placed it within the family.[18] In the APG III system, Strasburgeria is grouped with Ixerba to form the family Strasburgeriaceae in the rosid order Crossosomatales.[19]
More rarely, the genus Diegodendron was thought to be close to Strasburgeria and Ochnaceae. Molecular phylogenetic studies have strongly supported the inclusion of Diegodendron in the rosid order Malvales, and it is sometimes treated as a monospecific family therein.[20] Otherwise, it has been placed in Bixaceae, tho there is reason to suspect that it might be closer to Sphaerosepalaceae.[19]
All of the genera mentioned above, as well as Ochnaceae, were long regarded as anamalous taxa of uncertain affinity. All had been placed, at one time or another, with Ochnaceae, near Theaceae, a family now included in the basal asterid order Ericales.[19]
In 2012, an analysis of chloroplast DNA resolved Ochnaceae as sister to a group of five families known as the clusioids.[5] This result had only weak bootstrap support. The clusioids were once regarded as a group of four families,[21] but Clusiaceae was divided in 2009[6] and the name Calophyllaceae was resurrected for one of the resulting segregates.[1][22]
There are only a few morphological characters that unite the clusioids with Ochnaceae. The petal aestivation is often contort in the clusioids, and usually so in Ochnaceae. In both groups, the flowers usually bear numerous stamens, and in the ovary, the placentation is mostly axile. In the ovules, the nucellus is often thin, and the outer integument is usually thicker than the inner.[23]
Taxonomy
Ochnaceae is divided into three subfamilies: Medusagynoideae, Quiinoideae, and Ochnoideae.[4] A molecular phylogenetic study resolved Medusagynoideae and Quiinoideae as sister subfamilies, but this result had only weak statistical support.[5]
In both subfamilies, the flowers are polystemonous. Except for the genus Froesia, many or all of the flowers are unisexual. The anthers contain a massive septum between the thecae that persists after anther dehiscence. The styles radiate outward from the ovary. At anthesis, the ovary is sculpted with longitudinal ribs.[24]
Medusagynoideae consists of a single species, Medusagyne oppositifolia. It is endemic to the island of Mahé in the Seychelles.[25]
Quiinoideae comprises about 48 species in four genera: Froesia, Quiina, Touroulia, and Lacunaria. It is restricted to tropical America.[26] Froesia is distinct from the other three genera. Its flowers are always bisexual, and the fruit consists of three structures that resemble follicles, except that they are not quite entirely separate from each other.
The subfamily Ochnoideae was revised in 2014 as Ochnaceae sensu stricto.[3] In that treatment, 27 genera were described. An additional genus, Neckia, was resurrected in that same year, based on the results of a molecular phylogenetic study.[4] Amaral and Bittrich (2014) divided Ochnoideae into three tribes: Luxemburgieae, Sauvagesieae, and Ochneae. The genus Testulea was included in the tribe Sauvagesieae. No subtribes were recognized.
In 2014, a second reclassification of Ochnoideae was published, based on cladistic analysis of DNA sequences. In that paper, Schneider et alii divided Ochnoideae into four tribes: Testuleeae, Luxemburgieae, Sauvagesieae, and Ochneae. Their circumscription of the tribes was the same as that of Amaral and Bittrich (2014), except that Testulea was removed from Sauvagesieae into its own tribe, Testuleeae. Inclusion of Testulea in Sauvagesieae renders that tribe paraphyletic over Luxemburgieae.
In 1902, Philippe van Tieghem recognized the following eight families in what is now Ochnaceae sensu lato:
- Euthemidaceae Van Tiegh.
- Lophiraceae Endl.
- Luxemburgiaceae Van Tiegh.
- Medusagynaceae
- Quiinaceae
- Sauvagesiaceae Dum.
- Wallaceaceae Van Tiegh.
- Ochnaceae sensu stricto
These are now all treated as synonyms of Ochnaceae sensu lato.
References
- ^ a b c Angiosperm Phylogeny Group (2009). "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III" (PDF). Botanical Journal of the Linnean Society. 161 (2): 105–121. doi:10.1111/j.1095-8339.2009.00996.x. Retrieved 2013-07-06.
- ^ a b c Vernon H. Heywood, Richard K. Brummitt, Ole Seberg, and Alastair Culham. Flowering Plant Families of the World. Firefly Books: Ontario, Canada. (2007). ISBN 978-1-55407-206-4.
- ^ a b c d e f g h Maria do Carmo E. Amaral, and Volker Bittrich. 2014. "Ochnaceae". pages 253-268. doi:10.1007/978-3-642-39417-1_19 In: Klaus Kubitzki (editor). 2014. The Families and Genera of Vascular Plants volume XI. Springer-Verlag: Berlin, Heidelberg,, Germany. ISBN 978-3-642-39416-4 (print). ISBN 978-3-642-39417-1 (eBook). doi:10.1007/978-3-642-39417-1
- ^ a b c d e f g h i j Julio V. Schneider, Pulcherie Bissiengou, Maria do Carmo E. Amaral, Ali Tahir, Michael F. Fay, Marco Thines, Marc S.M. Sosef, Georg Zizka, and Lars W. Chatrou. 2014. "Phylogenetics, ancestral state reconstruction, and a new infrafamilial classification of the pantropical Ochnaceae (Medusagynaceae, Ochnaceae s.str., Quiinaceae) based on five DNA regions". Molecular Phylogenetics and Evolution 78:199-214. doi:10.1016/j.ympev.2014.05.018.
- ^ a b c Zhenxiang Xi, Brad R. Ruhfel, Hanno Schaefer, André M. Amorim, Manickam Sugumaran, Kenneth J. Wurdack, Peter K. Endress, Merran L. Matthews, Peter F. Stevens, Sarah Mathews, and Charles C. Davis. 2012. "Phylogenomics and a posteriori data partitioning resolve the Cretaceous angiosperm radiation Malpighiales". PNAS (Proceedings of the National Academy of Sciences U.S.A.) 109(43):17519-17524. doi:10.1073/pnas.1205818109. (See External links below).
- ^ a b Kenneth J. Wurdack and Charles C. Davis (2009), "Malpighiales phylogenetics: Gaining ground on one of the most recalcitrant clades in the angiosperm tree of life", American Journal of Botany, 96 (8): 1551–1570, doi:10.3732/ajb.0800207, PMID 21628300
- ^ Paul A. De Luca and Mario Vallejo-Marin. 2013. "What's the buzz about? The ecology and evolutionary significance of buzz pollination". Current Opinion in Plant Biology 16(4):429-435. doi:10.1016/j.pbi.2013.05.002.
- ^ Anthony Huxley, Mark Griffiths, and Margot Levy (1992). The New Royal Horticultural Society Dictionary of Gardening. The Macmillan Press,Limited: London. The Stockton Press: New York. ISBN 978-0-333-47494-5 (set).
- ^ Warren L. Wagner, Derral R. Herbst, and Sy H. Sohmer. Manual of the Flowering Plants of Hawaii, Revised Edition, 1999. Bishop Museum Press: Hololulu
- ^ David J. Mabberley. 2008. Mabberley's Plant-Book third edition (2008). Cambridge University Press: UK. ISBN 978-0-521-82071-4 (See External links below).
- ^ Peter K. Endress. 2011. "Evolutionary diversification of the flowers in angiosperms". American Journal of Botany 98(3):370-396. doi:10.3732/ajb.1000299. (See External links below).
- ^ Daniel Danehy, Peter Wilf, and Stefan A. Little. 2007. "Early Eocene macroflora from the red hot truck stop locality (Meridian, Mississippi, USA)". Palaeontologia Electronica 10(3):17A:31pages. (See External links below).
- ^ Susana Magallon, Khidir W. Hilu, and Dietmar Quandt. 2013. "Land plant evolutionary timeline: Gene effects are secondary to fossil constraints in relaxed clock estimation of age and substitution rates". American Journal of Botany 100(3):556-573. doi:10.3732/ajb.1200416. (See External links below).
- ^ The Plant List: Ochnaceae. (See External links below).
- ^ a b Julio V. Schneider, Ulf Swenson, Rosabelle Samuel, Tod Stuessy, and Georg Zizka. 2006. "Phylogenetics of Quiinaceae (Malpighiales): evidence from trnL-trnF sequence data and morphology". Plant Systematics and Evolution 257(3-4):189-203. doi:10.1007/s00606-005-0386-5.
- ^ a b Klaus Kubitzki. 2014. "Quiinaceae". pages 277-281. doi:10.1007/978-3-642-39417-1_22. In: Klaus Kubitzki (editor). 2014. The Families and Genera of Vascular Plants volume XI. Springer-Verlag: Berlin, Heidelberg,, Germany. ISBN 978-3-642-39416-4 (print). ISBN 978-3-642-39417-1 (eBook). doi:10.1007/978-3-642-39417-1
- ^ a b William C. Dickison and Klaus Kubitzki. 2014. "Medusagynaceae". pages 249-251. doi:10.1007/978-3-642-39417-1_18. In: Klaus Kubitzki (editor). 2014. The Families and Genera of Vascular Plants volume XI. Springer-Verlag: Berlin, Heidelberg,, Germany. ISBN 978-3-642-39416-4 (print). ISBN 978-3-642-39417-1 (eBook). doi:10.1007/978-3-642-39417-1
- ^ Armen L. Takhtajan (Takhtadzhian). Flowering Plants second edition (2009). Springer Science+Business Media. ISBN 978-1-4020-9608-2. eISBN 978-1-4020-9609-9 doi:10.1007/978-1-4020-9609-9. (See External links below).
- ^ a b c Peter F. Stevens (2001 onwards). "Ochnaceae" At: Angiosperm Phylogeny Website. At: Missouri Botanical Garden Website. (see External links below)
- ^ Clemens Bayer. 2003. Diegodendron. pages 175-177. In: Klaus Kubitzki (editor). 2003. The Families and Genera of Vascular Plants volume V. (volume editors: K. Kubitzki and C. Bayer). Springer-Verlag: Berlin, Heidelberg,, Germany. ISBN 978-3-540-42873-2 (print) ISBN 978-3-662-07255-4 (eBook). doi:10.1007/978-3-662-07255-4.
- ^ Peter F. Stevens. 2007. "Clusiaceae – Guttiferae". pages 48-66. In: Klaus Kubitzki (editor). 2007. The Families and Genera of Vascular Plants volume IX. (volume editors: K. Kubitzki in collaboration with C. Bayer and P.F. Stevens). Springer-Verlag: Berlin, Heidelberg,, Germany. ISBN 978-3-540-32214-6 () ISBN 978-3-540-32219-1 () doi:10.1007/978-3-540-32219-1.
- ^ Brad R. Ruhfel, Peter F. Stevens, and Charles C. Davis. 2013. "Combined morphological and molecular phylogeny of the clusioid clade (Malpighiales) and the placement of the ancient rosid macrofossil Paleoclusia". International Journal of Plant Sciences 174(6):910–936. doi:10.1086/670668.
- ^ Peter K. Endress, Charles C. Davis, and Merran L. Matthews. 2013. "Advances in the floral structural characterization of the major subclades of Malpighiales, one of the largest orders of flowering plants". Annals of Botany 111(5): 969–985. doi:10.1093/aob/mct056. (See External links below).
- ^ Merran L. Matthews, Maria do Carmo E. Amaral, and Peter K. Endress. 2012. "Comparative floral structure and systematics in Ochnaceae s.l. (Ochnaceae, Quiinaceae and Medusagynaceae; Malpighiales)". Botanical Journal of the Linnean Society 170(3):299-392. doi:10.1111/j.1095-8339.2012.01299.x.
- ^ William C. Dickison. 1990. "The morphology and relationships of Medusagyne (Medusagynaceae)". Plant Systematics and Evolution 171(1-4):27-55.
- ^ Julio V. Schneider and Georg Zizka. in press. "Quiinaceae". Flora Neotropica monograph ??.
External links
- Xi2012 At: Pubmed Central At: NCBI (National Center for Biotechnology Information)
- Cespedesia AndOchna In Mabberley's Plant-book
- Endress (2011) And land plant evolutionary timeline At: Archive At: American Journal of Botany
- Eocene macroflora from Mississippi (print) At: table of contents At: volume 10, issue 3 At: back issues by cover At: Browse Articles At: Palaeontologia Electronica/content
- Ochnaceae At: angiosperms At: Browse At: The Plant List
- Ochnaceae In Takhtajan 2009
- Ochnoideae distribution map AndOchnoideae genus list At Ochnoideae AtMalpighiales At Trees At: APweb At: Missouri Botanical Garden
- the clusioid clade and Paleoclusia
- floral structure in Malpighiales
{kind=link}
- Ochnaceae in L. Watson and M.J. Dallwitz (1992 onwards). The families of flowering plants: descriptions, illustrations, identification, information retrieval. http://delta-intkey.com
This Malpighiales article is a stub. You can help Wikipedia by expanding it. |