
Lepidodendron
| Lepidodendron | |
|---|---|

| |
| The strobilus of Lepidodendron | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Clade: | Lycophytes |
| Class: | Lycopodiopsida |
| Order: | †Lepidodendrales |
| Family: | †Lepidodendraceae |
| Genus: | †Lepidodendron Sternberg |
| Species | |
| |
Lepidodendron is an extinct genus of primitive, vascular, tree-like plants, also known as scale trees, related to the quillworts and lycopsids (club mosses). They were part of the coal forest flora. They sometimes reached heights of 50 metres (160 feet),[1] and the trunks were often over 1 m (3.3 ft) in diameter. They thrived during the Carboniferous Period (about 359.2 ± 2.5 Mya (million years ago) and were found until the Late Triassic, about 205 Mya) before going extinct.[2] Sometimes erroneously called "giant club mosses", the genus was actually more closely related to modern quillworts than to modern club mosses.
Etymology
The name Lepidodendron comes from the Greek λεπίς lepis, scale, and δένδρον dendron, tree.
Description and biology

Lepidodendron species were comparable in size to modern trees. The plants had tapering trunks as wide as 2 m (6.6 ft) at their base that rose to about 40 m (130 ft)[3] and even 50 m (160 ft),[1] arising from an underground system of horizontally spreading branches that were covered with many rootlets. Though the height of the trees make the plants similar to modern trees, the constant dichotomy of branches created a habit that contrasts with that of modern trees. At the ends of branches were oval-shaped cones that had a similar shape to modern cones of a spruce or fir.[4]
The stem of the trees had a unifacial vascular cambium, contrasting with the bifacial vascular cambium of modern trees. Though the bifacial cambium of modern trees produces both secondary phloem and xylem, the unifacial cambium of Lepidodendron trees produced only secondary xylem. As the trees aged, the wood produced by the unifacial cambium decreased towards the top of the plant such that terminal twigs resembled young Lepidodendron stems. The stems and branches of the trees contained little wood as compared to modern trees, with the majority of mature stems consisting of a massive cortical meristem. The near uniform growth of this cortical tissue indicates no difference in growth during changing seasons, and the absence of dormant buds further indicates the lack of seasonality in Lepidodendron species.[5] The outermost cortex of oldest stems developed into the bark-like lycopodiopsid periderm.[6] The bark of the trees was somewhat similar to that of Picea species, as leaf scars formed peg-like projections that stretched and tore as the bark stretched. To resist the bending force of wind, Lepidodendron trees depended on their outer bark rather than their vascular tissues, as compared to modern trees that rely mostly on their central mass of wood.[4]
The leaves of the trees were needle-like and were densely spiraled about young shoots, each possessing only a single vein. The leaves were similar to those of a fir in some species and similar to those of Pinus roxburghii in others, though in general the leaves of Lepidodendron species are indistinguishable from those of Sigillaria species. The decurrent leaves formed a cylindrical shell around branches. The leaves were only present on thin and young branches, indicating that though the trees were evergreen they did not retain their needles for as long as modern conifers. The leaf-cushions were fusiform and elongated, growing at most to a length of 8 cm (3.1 in) and a width of 2 cm (0.79 in). The middle of leaf-cushions were smooth, where leaf scars were created when an abscission layer cut a leaf from its base. Each leaf scar was composed of a central circular or triangular scar and two lateral scars that were smaller and oval-shaped. This central scar marks where the main vascular bundle of the leaf connected to the vascular system of the stem. This xylem bundle was composed only of primary trachea. The two outer scars mark the forked branches of a strand of vascular tissue that passed from the cortex of the stem into the leaf. This forked strand is sometimes referred to as the "parichnos". Surrounding this strand were parenchyma cells and occasionally thick-walled elements. Surrounding both conducting tissues was a broad sheath of transfusion tracheids. Below the leaf scar the leaf-cushion tapered to a basal position. In this tapering area circular impressions with fine pits were present. These impressions were continuous with the parichnos scars near the top of the tapering portion. This is because the impressions are formed by aerenchyma tissue that developed in closely with the parichnos. Above the leaf scar was a deep triangular impression known as the "ligular pit" for its similarities to the ligule of Isoetes. In some leaf-cushions a second depression was present above the ligular pit. Though its purpose is unclear, it has been suggested that the depression may mark the position of a sporangium. As the branch of a Lepidodendron tree grew the leaf-cushion only grew to a certain extent, past which the leaf-cushion stretched. This stretching widened the groove that separated the leaf-cushions, creating a broad, flat channel.[4]
Hyphae are occasionally present in the tissues of Lepidodendron trees, indicating a susceptibility to fungal parasites.[4]
Different fossil genera have been described to name the various levels of decay in Lepidodendron bark fossils. The name Bergeria describes stems that have lost their epidermises, Aspidiariu is used when cushions have been removed by deep decay, and Knorria is used when the leaf cushions and the majority of cortical tissues has decayed, with a shallow "fluted" surface remaining. However, it has been suggested that these are more likely growth forms than preserved bark types, as entire fossilized trunks have been discovered with dissimilar forms; if decay is assumed to be constant throughout the trunk, then different forms indicate growth rather than levels of decay. It is likely that the trunk of Lepidodendron trees were subject to the growth forms Knorria, Aspidiaria, and Bergeria progressing up the trunk, respectively.[7]
Reproduction
Lepidodendron species had a life cycle of 10 to 15 years composed of a growth cycle, in which the trees grew to a predetermined height, and a subsequent reproductive cycle, in which the trees produced reproductive organs, after which the trees died, similar to the life cycle of a Mauna Kea silversword.[6]
Rather than reproduce with seeds, Lepidodendron trees reproduced with spores. The spores were stored in sporangia situated on fertile stems that grew on or near the main trunk. The fertile stems grew together in cone-like structures that clustered at the tips of branches.[8]
Distribution
The lack of growth rings and of dormant buds indicate no seasonal growth patterns and modern plants with similar characteristics tend to grow in tropical conditions, but Lepidodendron species were distributed throughout subtropical conditions. The trees inhabited an extensive area compared to tropical flora of the same time period, with trees growing as far north as Spitsbergen and as far south as South America, in a latitudinal range of 120°.[5]
Decline and extinction
By the Mesozoic era, the giant lycopsids had died out and were replaced by conifers as well as smaller quillworts.[2] This may have been the result of competition from the emerging woody gymnosperms. Lepidodendron is one of the more common plant fossils found in Pennsylvanian-age (Late Carboniferous) rocks. They are closely related to other extinct lycopsid genera, Sigillaria and Lepidendropsis.
This section needs expansion. You can help by adding to it. (May 2012) |
In popular culture
In the 19th century, due to the reptilian look of the diamond-shaped leaf scar pattern, petrified trunks of Lepidodendron were frequently exhibited at fairgrounds by amateurs as giant fossil lizards or snakes.[9] Inversely, the fossil of a temnospondyl amphibian collected in a Triassic formation in Tasmania was described in 1885 as the cone of an ancient plant, with affinities to this genus, as Lepidostrobus muelleri.[10][11]
Gallery
-
 External mold of Lepidodendron from the Upper Carboniferous of Ohio
External mold of Lepidodendron from the Upper Carboniferous of Ohio -
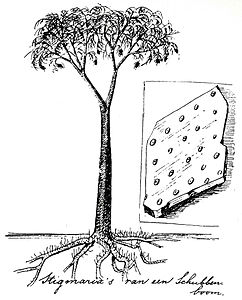 Reconstruction of Lepidodendron by Dutch conservationist Eli Heimans (1911)
Reconstruction of Lepidodendron by Dutch conservationist Eli Heimans (1911) -
 Lepidodendron on display at the State Museum of Pennsylvania
Lepidodendron on display at the State Museum of Pennsylvania



See also
References
- ^ a b V. V. Alekhin (1961). Geografiia rastenii s osnovani botaniki (Geography of plants and basics of botany). Gos. nauchno-pedagog. izd-vo. p. 167. Retrieved 2020-10-05.
- ^ a b Sahney, S., Benton, M.J. & Falcon-Lang, H.J. (2010). "Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica". Geology. 38 (12): 1079–1082. doi:10.1130/G31182.1.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ A. V. Lopatin (2012). Палеонтологический музей имени Ю.А. Орлова (The Orlov Museum of Paleontology). Moscow: PIN RAN. p. 56. ISBN 978-5-903825-14-1. Retrieved 2020-10-05.
- ^ a b c d Seward, Albert Charles (1898). Fossil plants: for students of botany and geology. Vol. 1. Cambridge University Press. pp. 93–192.
- ^ a b Vulf, Evgenii Vladimirovich and Brissenden, Elizabeth (1943). An introduction to historical plant geography. Chronica Botanica Company. pp. 176–177.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ^ a b Karl J. Niklas (1997). The Evolutionary Biology of Plants (illustrated ed.). University of Chicago Press. p. 321. ISBN 9780226580838.
- ^ Thomas, B.A. and Watson, Joan (1976). "A rediscovered 114-foot Lepidodendron from Bolton, Lancashire". Geological Journal. 11 (1). Wiley Online Library: 15–20. doi:10.1002/gj.3350110102.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ John Adam Dorr, Donald F. Eschman (1970). Geology of Michigan (illustrated ed.). University of Michigan Press. p. 429. ISBN 9780472082803.
- ^ Robert Logan Jack (1886). Handbook of Queensland geology. Geological Survey of Queensland. Warwick and Sapsford. p. 28.
- ^ Johnston, R.M. (1885). "Discovery of a cone, probably of a species of Lepidostrobus, in the sandstones of Campania". Papers and Proceedings of the Royal Society of Tasmania. 1884. The Society.: 225.
- ^ Rozefelds, A.C.; Warren, A. (September 2011). "Lepidostrobus muelleri Johnston is a skull fragment of a temnospondyl amphibian". Alcheringa: An Australasian Journal of Palaeontology. 35 (3): 459–462. doi:10.1080/03115518.2011.527163. S2CID 83845584.
Further reading
- Davis, Paul; Kenrick, Paul (2004). Fossil Plants. Washington, DC: Smithsonian Books. ISBN 1-58834-181-X.
- Morran, Robin C. (2004). A Natural History of Ferns. Portland: Timber Press. ISBN 0-88192-667-1.
- "Plant fossils of the British Coal Measures" by Christopher J.Cleal and Barry A.Thomas, publ. The Palaeontological Association, London, 1994, 222 pages, ISBN 0-901702-53-6
- J. M. Anderson and H. M. Anderson. 1985. Palaeoflora of Southern Africa. Prodromus of South African Megafloras Devonian to Lower Cretaceous 1-423
- Lycophytes
- Prehistoric trees
- Pennsylvanian plants
- Carboniferous life of North America
- Fossils of Georgia (U.S. state)
- Paleozoic life of New Brunswick
- Paleozoic life of Newfoundland and Labrador
- Paleozoic life of the Northwest Territories
- Paleozoic life of Nova Scotia
- Paleozoic life of Nunavut
- Paleozoic life of Quebec
- Permian Africa
- Fossils of South Africa
- Paleozoic life of Oceania
- Permian life of Australia
- Fossils of Australia
- Paleozoic life of Asia
- Permian China
- Fossils of China
- Fossils of Indonesia
- Fossils of North Korea
- Fossils of Oman
- Fossils of South Korea
- Paleozoic life of Europe
- Fossils of Italy
- Fossil taxa described in 1820
- Lycophyte genera