
POLD1





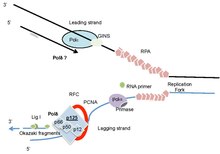
The gene polymerase delta 1 (POLD1) encodes the large, POLD1/p125, catalytic subunit of the DNA polymerase delta (Polδ) complex.[5][6] The Polδ enzyme is responsible for synthesizing the lagging strand of DNA, and has also been implicated in some activities at the leading strand (Figure 1). The POLD1/p125 subunit encodes both DNA polymerizing and exonuclease domains, which provide the protein an important second function in proofreading to ensure replication accuracy during DNA synthesis, and in a number of types of replication-linked DNA repair following DNA damage. Germline mutations impairing activity of POLD1 have been implicated in several types of hereditary cancer, in some sporadic cancers, and in a developmental syndrome of premature aging, Mandibular hypoplasia, Deafness, and Progeroid features and Lipodystrophy (MDPL/MDP syndrome). Studies of POLD1 emphasize the importance of maintaining genomic stability to limit tumorigenesis. It is currently unclear whether the enhanced tumorigenesis associated with POLD1 defects is the result of increased base substitutions or due to fork collapse and production of DNA double strand breaks (DSBs).[6][7] Recent reviews have addressed important functions of POLD1 and Polδ.[6][7]
Discovery
The first DNA polymerase, DNA polymerase I, was discovered by Arthur Kornberg and his colleagues in 1956,[8] reviewed in.[9] In 1976, Byrnes et al. discovered a third DNA polymerase activity in mammalian cells that was called polymerase delta (δ).[10] It was purified from rabbit erythroid hyperplastic bone marrow and described as a DNA polymerase that possessed an intrinsic 3’ to 5’ exonuclease activity. A 3’-5’ exonuclease proofreading function for DNA polymerases (E. coli) had first been described 4 years earlier by Kornberg and Brutlag,[11] reviewed in.[12] The human DNA Polδ is a heterotetramer. The four subunits are: (POLD1/ p125), (POLD3/ p66), (POLD2/ p50) and (POLD4/ p12), with the alternative names reflecting the molecular weights expressed in kilodaltons (kDa). The polymerase catalytic subunit was identified as the 125 kDa polypeptide by activity staining in 1991.[13] Several groups independently cloned the human and murine POLD1 cDNAs.[5][14][15] Following its purification from various sources including calf thymus, human placenta, and HeLa cells,[16][17][18][19][20] its activity was implicated in DNA repair.[21][22]
Gene
Polymerase (DNA) delta 1, catalytic subunit and POLD1 are the name and gene symbol approved by the Human Genome Organization (HUGO) Gene Nomenclature Committee (HGNC).[23] POLD1 is also known as CDC2, MDPL, POLD, and CRCS10), is ~34 kb long and its cytogenetic location is chromosome 19[24] q13.33.[25] The precise location, in the GRCh38.p2 assembly, is from base pair 50,384,290 to base pair 50,418,018 on chromosome 19.[26] The mouse orthologue maps to mouse chromosome 7.[27] In humans, the major POLD1 transcript (NM_002691.3) contains 27 exons and translates into the 1107 amino acids of the p125 or A subunit. A longer isoform has been reported with a 26 amino acid in-frame insertion after amino acid 592 (NP_001295561.1). A pseudogene (LOC100422453) has been reported on the long arm of chromosome 6.[26] Table 1 provides gene names and chromosomal locations for the various subunits of Polδ in humans, mice, budding yeast (S. cerevisiae) and fission yeast (S. pombe).
The POLD1 gene promoter is regulated via the cell cycle machinery and mRNA expression of POLD1 reaches a peak in late G1/S phase during DNA replication.[28] The POLD1 promoter is G/C-rich and has no TATA box. The transcription of this GC box-containing promoter is regulated by Sp1 and Sp1-related transcription factors such as Sp3, with their binding mediated via 11-bp repeat binding sequences.[29][30] The POLD1 promoter contains an E2F-like sequence located near the major transcription start site.[30] Another regulatory element, the cell cycle element/cell cycle genes homology region (CDE/CHR), located downstream of the start site is important for POLD1 transcription in G2/M phase by E2F1 and p21 proteins.[31][32] P53 regulates POLD1 transcription by indirect p21-dependent activation of a p53-p21-DREAM-CDE/CHR pathway.[33] One study has reported that the p53 tumor suppressor protein competes with Sp1 for binding to the POLD1 promoter.[29] A microRNA (miR), miR-155, downregulates POLD1 indirectly by suppressing the transcription factor FOXO3a,[34] which has putative binding sites in the POLD1 promoter (RTMAAYA; response element).[35]
Protein
POLD1/p125 has a common B-family fold, similar to other DNA polymerases (Polα and ε).[37] Human POLD1/p125 has a putative nuclear localization signal at the N-terminal end (residues 4-19).[24] Residues 304-533 contain the exonuclease domain (Figure 2) while residues 579-974 contain the polymerase domain. The exonuclease domain is a DEDDy-type DnaQ-like domain common to the B-DNA polymerase family.[38] This domain has a beta hairpin structure that helps in switching between the polymerase and exonuclease active sites in case of nucleotide misincorporation.
Motifs A and C, which are the most conserved of the polymerase domain. These have 2 catalytic aspartates, in motif A (DXXLYPS, D602) and motif C (DTDS, D757) that bind calcium at the active site. Motif A has 11 amino acids that are important in nucleotide incorporation and formation of the phosphodiester bond.
Tyrosine Y701 functions similarly to tyrosine Y567 in the RB69 bacteriophage orthologue as the sugar steric gate that prevents ribonucleotide incorporation.[39] An LXCXE motif (711 to 715) mediates binding to pRB during the G1 phase of cell cycle.[40] The polymerase domain also has a highly conserved KKRY motif (residues 806 to 809) which is important for the binding and catalytic function.[41] POLD1 can be targeted to the nucleolus upon acidification via a nucleolar detention sequence (NoDS) motif represented by small sequence motifs dispersed throughout the protein coding region.[42][43][44] The C-terminal domain has two conserved cysteine-rich metal-binding motifs (CysA and CysB) (from 1012 and 1083) required for Proliferating Cell Nuclear Antigen (PCNA) binding and recruitment of accessory subunits respectively.[45] CysB coordinates an [4Fe-4S] cluster added through Cytosolic Iron-sulfur protein Assembly (CIA), which requires the function of the mitochondrial Iron Sulfur Cluster (ISC) assembly machinery.[46] The maturation process is mediated by the core targeting complex CIA1-CIA2B/FAM96B-MMS19, which interacts with the apoprotein to ensure specific Fe-S cluster insertion.[47][48]

Binding and association studies have shown that POLD2 is tightly associated with POLD1; POLD3 and POLD2 interact with each other and POLD4 interacts with both POLD1 and POLD2.[51][52] Polδ heterotetramer reconstituted by coexpression of subunits in Sf9 cells had properties were similar to Polδ purified from the calf thymus, and the complete holoenzyme was very strongly stimulated by PCNA.[53] Numerous studies have shown that while POLD1 possesses both the polymerase and the 3’-5’ exonuclease proofreading activity, the other subunits increase these activities, DNA binding abilities, and functionally important interactions with PCNA and its clamp loader Replication Factor C (RFC). The DNA Polδ holoenzyme is often considered to include PCNA and RFC as well as the four subunits of the polymerase complex (Figure 1).
A number of other studies and screens have identified additional interaction partners relevant to functions in DNA replication and repair. Figure 3 shows a matrix of established and putative interactions during replication and repair which can be further accessed through[54] and.[55] A website at Vanderbilt University provides additional interaction on important POLD1 protein structure and various classes of gene and protein interaction, based on criteria such as co-occurrence in a complex, direct physical interaction, regulatory relationship, and co-expression.[56]
| Polymerase delta subunits |
Protein name in human |
Homo sapiens | Mus musculus | Saccharomyces cerevisiae | Schizosaccharomyces pombe |
|---|---|---|---|---|---|
| A (catalytic) | p125 | POLD1-Chr 19q13.3 | Pold1-Chr 7B4 | POL3-Chr IV | cdc6-Chr II |
| B (accessory) | p50 | POLD2-Chr 7p13 | Pold2-Chr 11A2 | POL31-Chr X | cdc1-Chr I |
| C (accessory) | p66 | POLD3-Chr 11q14 | Pold3-Chr 7F1 | POL32-Chr X | cdc27-Chr II |
| D (accessory) | p12 | POLD4-Chr 11q13 | Pold4-Chr 19A | - | cdm1-Chr II |
| Table 1: Gene names and chromosomal locations for the various subunits of polymerase delta in human, mouse, budding and fission yeast. | |||||
Expression and regulation

The POLD1/P125 protein is expressed ubiquitously across a panel of human tissues with high levels in the heart and lung tissues.[60] The subcellular localization of POLD1/p125 is predominantly in the nucleus and nucleoplasm.[61]
A reduction in POLD1/p125 has been observed in senescent human skin fibroblasts and in lymphocytes from an elderly population.[62][63] POLD1/p125 expression is epigenetically regulated in response to DNA damage.[64] Other studies have also shown that POLD1/p125 expression is regulated by miR-155,[34] p53[29] and by the long non-coding RNA, PVT1.[65] In the presence of DNA damage or replication stress (UV light, methyl methanesulfonate, hydroxyurea or aphidicolin), the POLD4/p12 subunit is rapidly degraded. The catalytic activities of p125 are different whether it is in the heterotetramer (Polδ4, with p12 [66][67]) or in the heterotrimer (Polδ3, without p12).[68] The production of the heterotrimer depends on p12 degradation by the E3 ligase RNF8, a protein involved in DSBs repair and possibly homologous recombination (HR).[69] In addition, the E3 ligase CRL4Cdt2 can degrade POLD4/p12 during normal DNA replication and in the presence of DNA damage.[70] POLD4/p12 can also be degraded by the protease µ-calpain, that is involved in calcium-triggered apoptosis.[71][72]
POLD1/p125 has a NoDS domain that regulates transport to the nucleolus in response to acidosis.[44] Nucleolar transport requires a direct interaction between the p50 subunit and the WRN protein.[73] During DNA damage response, WRN moves out of the nucleolus and thereby releases Polδ.[74][75] POLD1/p125 has also been shown to interact with PDIP46/SKAR[76] and LMO2.[77][78]
Function
DNA replication
DNA replication is a highly organized process that involves many enzymes and proteins, including several DNA polymerases. The major replicative activity in S phase of cell cycle depends on three DNA polymerases - Polymerase alpha (Polα), Polymerase delta (Polδ), and Polymerase epsilon (Polε). After initiation of DNA synthesis by Polα, Polδ or Polε execute lagging and leading strand synthesis, respectively.[79] These polymerases maintain a very high fidelity, which is ensured by Watson-Crick base pairing and 3'-exonuclease (or the proofreading) activity.[80] Recent studies have contended that Polδ may synthesize the leading strand.[80][81][82][83][84] How these polymerases function, in relationship with other factors involved in replication, is of great interest as it likely explains the mutational landscape that they produce when defective. Maintenance of replication fidelity is a fine balance between the unique errors by polymerases δ and ε,[85] the equilibrium between proofreading and MMR, and distinction in ribonucleotide processing between the two strands.[36] Extensive studies in yeast models have shown that mutations in the exonuclease domain of Polδ and Polε homologues can cause a mutator phenotype, reviewed in.[86] The single stranded (ss) DNA synthesized during lagging strand synthesis can be targeted by ss-DNA damaging agents as well as is a selective target for APOBEC mutations.[87] DNA-binding proteins that rapidly reassociate post-replication prevent Polδ from repairing errors produced by Polα in the mature lagging strand.[88] Yeast studies have shown that Polδ can proofread Polε errors on the leading strand.[89]
DNA Repair
POLD1 activity contributes to multiple evolutionarily conserved DNA repair processes, including Mismatch repair (MMR), Translesion synthesis (TLS), Base excision repair (BER), Nucleotide Excision repair (NER) and double-strand break (DSB) repair.[6] POLD1 mediates the post-incision steps in BER, NER and MMR.[6] Polδ interacts with the MMR machinery to support post-replication proofreading of newly synthesized DNA,[90] with cells bearing mutations that inactivate POLD1 and MMR components experiencing elevated mutation rates.[91][92] As noted above, a Polδ heterotrimer (Polδ3) becomes the dominant oligomeric form of POLD1 and is active during the presence of DNA damage. Polδ3 is less error-prone than (Polδ4), and can discriminate better between mismatched pairs, associated with better proofreading activity: however, it has reduced ability to bypass some base lesions.[74][93] Instead, Polδ polymerase switching to the specialized polymerase zeta (Polζ) is important for TLS as the substitution of p125 for the Polζ catalytic subunit, p353, permits better bypass activity.[6] In this process, the highly conserved C-terminal domain (CTD) of POLD1/p125 interacts with the CTD domain of Polζ, and the iron clusters within each CTD mediate interactions involving binding to POLD2 that permit polymerase switching during TLS.[94] Some recent studies suggest that a switch from Polδ to Pol lambda (λ) also supports the TLS and repair of oxidative DNA damage like 7,8-Dihydro-8-oxoguanine lesions.[95]
Depletion of POLD1 can halt cell cycle at G1 and G2/M phases in human cells.[96] Cell cycle block in these phases typically indicates presence of DNA damage and activation of DNA damage checkpoints. POLD1 depleted cells are sensitive to inhibition of DNA damage checkpoint kinases ATR and CHK1.[97] In S. pombe, HR mechanisms could restart stalled replication forks by utilizing Polδ strand synthesis activity, but such nonallelic HR-mediated restart is very error prone potentially leading to increased genomic instability.[98] Polδ structurally and functionally interacts with the WRN protein, and WRN recruits Polδ to the nucleolus.[73] The WRN gene is mutated in Werner syndrome (an autosomal recessive disorder) leading to accelerated aging and increased genetic instability. The interaction with WRN increases the processivity of Polδ in a PCNA-independent manner.[99] Through these interactions WRN directly impacts DNA replication-repair and assists in Polδ-mediated synthesis.
Clinical significance
Cancer
DNA repair proteins have been shown to be important in human diseases including cancer. For example, germline mutations in DNA repair proteins involved in MMR (MSH2, MLH1, MSH6, and PMS2) have been described in Lynch syndrome (LS), which is characterized by the presence of microsatellite instability (MSI).[100] More recently, germline mutations have been reported in the exonuclease domains of POLD1 and POLE, the catalytic subunit of Polε. These mutations are associated with oligo-adenomatous polyposis, early-onset colorectal cancer (CRC), endometrial cancer (EDMC), breast cancer, and brain tumors.([101][102][103][104][105] reviewed in[7]) Most of the reported POLD1 mutations linked to cancer are present in the exonuclease domain.[7][101][102][106][107][108] In contrast to LS, the POLD1 mutated tumors are microsatellite stable. Some data suggests the idea that POLD1 tumors are associated with driver mutations in genes including APC and KRAS.[101] The POLD1 missense mutation p. S478N, in the exonuclease domain, has been validated as damaging and pathogenic.[101] Other POLD1 variants have been clinically identified which have been predicted to be damaging and are currently under further investigation (e.g., p. D316H, p. D316G, p. R409W, p. L474P and p. P327L).[102][103][104]
In pediatric patients, double hit mutations in POLD1 or POLE and biallelic mismatch repair deficiency (bMMRD), leads to ultra-hypermutated tumor phenotypes.[109][110][111] Such phenotypes as ultra-hypermutation in tumors may indicate better response to newer cancer therapeutics in development, although this needs direct evaluation for POLD1.[112][113][114][115][116][117] Bouffet et al. report two siblings with bMMRD- glioblastoma multiforme who have somatic mutations in POLE (P436H in one, S461P in the other), and showed a durable response to a clinical trial with the anti-programmed death-1 inhibitor nivolumab. POLD1 mutations have been studied in cell lines [118][119][120][121] and mouse models. For example, a homozygous Polδ mutation in mice that disrupts enzymatic function leads to highly elevated cancer incidence.[122]
MDPL
Damaging mutations in POLD1 have also been observed in patients with a syndrome known as mandibular hypoplasia, deafness, and progeroid features with lipodystrophy (MDPL/MDP) syndrome (#615381 in the Online Mendelian Inheritance in Man (OMIM) database).[60][123][124] This is a very rare syndrome, and few studies describing mutations have been reported. The mutations that have been observed are in the regions that affect the exonuclease domain and polymerase domains.[60][123] Five unrelated de novo cases have been described with the same heterozygous variant, c.1812_1814delCTC p.Ser605del (rs398122386). S605 is in the highly conserved motif A of the polymerase active site. This variant does not inhibit the DNA binding activity but impacts catalysis. Another variant has been reported in a separate patient (p.R507C).[123] This variant is located in the highly conserved ExoIII domain and has not been completely characterized as yet.
Interestingly, POLD1 Ser605del and R507C variants have also been identified in a subset of patients with atypical Werner’s syndrome (AWS). After molecular testing, these patients were reclassified as MDPL/MDP patients. MDPL/MDP, AWS and Werner’s syndrome all present with progeria.[125] A first example of germline transmission was observed in a mother and son with the Ser605del mutation.[126] Recently, two independent studies identified patients with the same homozygous splice variant in POLE1, the catalytic subunit of Polε. One presented with a phenotype of facial dysmorphism, immunodeficiency, livedo, and short stature (also knowns as the FILS syndrome).[127] The second one presented with more severe symptoms.[128] These cases join a growing number of developmental defects associated with inherited mutations targeting the function of polymerase genes.
In this context, it may be interesting that age-dependent downregulation of POLD1 has been observed.[63] although no clinical significance has been associated with this phenotype as yet. Studies are also underway to understand if there is a relation between these pathologies or these mutations and a predisposition to cancer. Currently proposed mechanisms by which POLD1 defects are pathogenic focus on the idea of replication defects leading to genomic instability and checkpoint activation, ultimately leading to cell death or cellular senescence. Alternatively, Polδ is associated with lamins and the nuclear envelope during G1/S arrest or early S phase; mutations in lamins cause nuclear envelope-related lipodystrophies with phenotypes similar to MDPL/MDP and Werner’s syndrome.[129]
Cancer risk assessment and commercial testing
The hereditary colorectal cancers (CRCs) associated with mutations in the proofreading ability of POLD1 and POLE are sometimes termed as “polymerase proofreading associated polyposis” (PPAP), (although at least one study has identified POLD1 mutations associated with non-polyposis CRC).[101][102][104][106][107] POLD1 mutations have also been associated with an increased cancer predisposition of endometrial cancer.[101][104][105] A recent study has suggested guidelines for genetic testing for POLD1 mutations which include: 1) Occurrence of 20-100 adenomas, and 2) Family history that meets the Amsterdam II criteria for colorectal and endometrial cancers.[103] Current clinical testing guidelines for families with mutations in POLD1/POLE include colonoscopies (every 1–2 years), gastroduodenoscopies (every 3 years) starting early (20-25), possibility for brain tumors and endometrial cancer screening (beginning at 40 for female carriers).[103] Currently studies are underway to determine the exact cancer risk from specific POLD1 mutations. Current data suggest that mutations in this gene are highly penetrant. Another recent study showed that mutations affecting Polδ and Polε mutations can co-occur along with MMR mutations.[110] This suggests panel gene testing should include MMR and Pol genes even in patients with MSI.
There are several options for commercial diagnostic testing for mutations in POLD1.[130] Genetic testing typically includes POLD1 coding exons (26) and at least 20 bases into the adjacent non-coding regions. For families with known mutations, single site testing is also available to confirm the presence of a mutation.[130] The availability of these genetic tests has opened up new possibilities for cancers previously classified as genetically undefined colorectal cancers or colorectal cancer type “X”.[105] Resources for clinical testing for MDPL/MDP have also been developed.[131]
References
- ^ a b c GRCh38: Ensembl release 89: ENSG00000062822 – Ensembl, May 2017
- ^ a b c GRCm38: Ensembl release 89: ENSMUSG00000038644 – Ensembl, May 2017
- ^ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ a b Chung DW, Zhang JA, Tan CK, Davie EW, So AG, Downey KM (December 1991). "Primary structure of the catalytic subunit of human DNA polymerase delta and chromosomal location of the gene". Proceedings of the National Academy of Sciences of the United States of America. 88 (24): 11197–201. doi:10.1073/pnas.88.24.11197. PMC 53101. PMID 1722322.
- ^ a b c d e f Prindle MJ, Loeb LA (December 2012). "DNA polymerase delta in DNA replication and genome maintenance". Environmental and Molecular Mutagenesis. 53 (9): 666–82. doi:10.1002/em.21745. PMC 3694620. PMID 23065663.
- ^ a b c d Rayner E, van Gool IC, Palles C, Kearsey SE, Bosse T, Tomlinson I, Church DN (January 2016). "A panoply of errors: polymerase proofreading domain mutations in cancer". Nature Reviews. Cancer. 16 (2): 71–81. doi:10.1038/nrc.2015.12. PMID 26822575.
- ^ Kornberg A, Kornberg SR, Simms ES (April 1956). "Metaphosphate synthesis by an enzyme from Escherichia coli". Biochimica et Biophysica Acta. 20 (1): 215–27. doi:10.1016/0006-3002(56)90280-3. PMID 13315368.
- ^ Friedberg EC (February 2006). "The eureka enzyme: the discovery of DNA polymerase". Nature Reviews Molecular Cell Biology. 7 (2): 143–7. doi:10.1038/nrm1787. PMID 16493419.
- ^ Byrnes JJ, Downey KM, Black VL, So AG (June 1976). "A new mammalian DNA polymerase with 3' to 5' exonuclease activity: DNA polymerase delta". Biochemistry. 15 (13): 2817–23. doi:10.1021/bi00658a018. PMID 949478.
- ^ "Enzymatic synthesis of deoxyribonucleic acid. XXXVI. A proofreading function for the 3 5′ exonuclease activity in deoxyribonucleic acid polymerase". ResearchGate. Retrieved 2016-04-25.
- ^ Reha-Krantz LJ (May 2010). "DNA polymerase proofreading: Multiple roles maintain genome stability". Biochimica et Biophysica Acta. 1804 (5): 1049–63. doi:10.1016/j.bbapap.2009.06.012. PMID 19545649.
- ^ Lee MY, Jiang YQ, Zhang SJ, Toomey NL (February 1991). "Characterization of human DNA polymerase delta and its immunochemical relationships with DNA polymerase alpha and epsilon". The Journal of Biological Chemistry. 266 (4): 2423–9. PMID 1703528.
- ^ Yang CL, Chang LS, Zhang P, Hao H, Zhu L, Toomey NL, Lee MY (February 1992). "Molecular cloning of the cDNA for the catalytic subunit of human DNA polymerase delta". Nucleic Acids Research. 20 (4): 735–45. doi:10.1093/nar/20.4.735. PMC 312012. PMID 1542570.
- ^ Cullmann G, Hindges R, Berchtold MW, Hübscher U (December 1993). "Cloning of a mouse cDNA encoding DNA polymerase delta: refinement of the homology boxes". Gene. 134 (2): 191–200. doi:10.1016/0378-1119(93)90093-i. PMID 8262377.
- ^ Lee MY, Tan CK, So AG, Downey KM (May 1980). "Purification of deoxyribonucleic acid polymerase delta from calf thymus: partial characterization of physical properties". Biochemistry. 19 (10): 2096–101. doi:10.1021/bi00551a015. PMID 7378348.
- ^ Lee MY, Tan CK, Downey KM, So AG (April 1984). "Further studies on calf thymus DNA polymerase delta purified to homogeneity by a new procedure". Biochemistry. 23 (9): 1906–13. doi:10.1021/bi00304a003. PMID 6426510.
- ^ Crute JJ, Wahl AF, Bambara RA (January 1986). "Purification and characterization of two new high molecular weight forms of DNA polymerase delta". Biochemistry. 25 (1): 26–36. doi:10.1021/bi00349a005. PMID 3954990.
- ^ Wahl AF, Crute JJ, Sabatino RD, Bodner JB, Marraccino RL, Harwell LW, Lord EM, Bambara RA (December 1986). "Properties of two forms of DNA polymerase delta from calf thymus". Biochemistry. 25 (24): 7821–7. doi:10.1021/bi00372a006. PMID 3099836.
- ^ Lee MY, Toomey NL (February 1987). "Human placental DNA polymerase delta: identification of a 170-kilodalton polypeptide by activity staining and immunoblotting". Biochemistry. 26 (4): 1076–85. doi:10.1021/bi00378a014. PMID 2436659.
- ^ Dresler SL, Kimbro KS (May 1987). "2',3'-Dideoxythymidine 5'-triphosphate inhibition of DNA replication and ultraviolet-induced DNA repair synthesis in human cells: evidence for involvement of DNA polymerase delta". Biochemistry. 26 (10): 2664–8. doi:10.1021/bi00384a002. PMID 3606985.
- ^ Nishida C, Reinhard P, Linn S (January 1988). "DNA repair synthesis in human fibroblasts requires DNA polymerase delta". The Journal of Biological Chemistry. 263 (1): 501–10. PMID 3335506.
- ^ "HGNC database of human gene names | HUGO Gene Nomenclature Committee". www.genenames.org. Retrieved 2016-04-25.
- ^ a b Chung DW, Zhang JA, Tan CK, Davie EW, So AG, Downey KM (December 1991). "Primary structure of the catalytic subunit of human DNA polymerase delta and chromosomal location of the gene". Proceedings of the National Academy of Sciences of the United States of America. 88 (24): 11197–201. doi:10.1073/pnas.88.24.11197. PMC 53101. PMID 1722322.
- ^ Kemper RR, Ahn ER, Zhang P, Lee MY, Rabin M (September 1992). "Human DNA polymerase delta gene maps to region 19q13.3-q13.4 by in situ hybridization". Genomics. 14 (1): 205–6. doi:10.1016/s0888-7543(05)80311-8. PMID 1427831.
- ^ a b "POLD1 polymerase (DNA) delta 1, catalytic subunit [Homo sapiens (human)] - Gene - NCBI". www.ncbi.nlm.nih.gov. Retrieved 2016-04-25.
- ^ Goldsby RE, Singh M, Preston BD (January 1998). "Mouse DNA polymerase delta gene (Pold1) maps to chromosome 7". Mammalian Genome. 9 (1): 92–3. doi:10.1007/s003359900693. PMID 9434960.
- ^ Mjelle R, Hegre SA, Aas PA, Slupphaug G, Drabløs F, Saetrom P, Krokan HE (June 2015). "Cell cycle regulation of human DNA repair and chromatin remodeling genes". DNA Repair. 30: 53–67. doi:10.1016/j.dnarep.2015.03.007. PMID 25881042.
- ^ a b c Li B, Lee MY (August 2001). "Transcriptional regulation of the human DNA polymerase delta catalytic subunit gene POLD1 by p53 tumor suppressor and Sp1". The Journal of Biological Chemistry. 276 (32): 29729–39. doi:10.1074/jbc.M101167200. PMID 11375983.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b Zhao L, Chang LS (February 1997). "The human POLD1 gene. Identification of an upstream activator sequence, activation by Sp1 and Sp3, and cell cycle regulation". The Journal of Biological Chemistry. 272 (8): 4869–82. doi:10.1074/jbc.272.8.4869. PMID 9030545.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Müller GA, Wintsche A, Stangner K, Prohaska SJ, Stadler PF, Engeland K (2014-01-01). "The CHR site: definition and genome-wide identification of a cell cycle transcriptional element". Nucleic Acids Research. 42 (16): 10331–50. doi:10.1093/nar/gku696. PMC 4176359. PMID 25106871.
- ^ Song N, Zhu X, Shi L, An J, Wu Y, Sang J (June 2009). "Identification and functional analysis of a CDE/CHR element in the POLD1 promoter". Science in China. Series C, Life Sciences / Chinese Academy of Sciences. 52 (6): 551–9. doi:10.1007/s11427-009-0077-5. PMID 19557333.
- ^ Fischer M, Quaas M, Steiner L, Engeland K (January 2016). "The p53-p21-DREAM-CDE/CHR pathway regulates G2/M cell cycle genes". Nucleic Acids Research. 44 (1): 164–74. doi:10.1093/nar/gkv927. PMC 4705690. PMID 26384566.
- ^ a b Czochor JR, Sulkowski P, Glazer PM (April 2016). "miR-155 Overexpression Promotes Genomic Instability by Reducing High-fidelity Polymerase Delta Expression and Activating Error-Prone DSB Repair". Molecular Cancer Research. 14 (4): 363–73. doi:10.1158/1541-7786.MCR-15-0399. PMID 26850462.
- ^ Chen X, Ji Z, Webber A, Sharrocks AD (February 2016). "Genome-wide binding studies reveal DNA binding specificity mechanisms and functional interplay amongst Forkhead transcription factors". Nucleic Acids Research. 44 (4): 1566–78. doi:10.1093/nar/gkv1120. PMC 4770209. PMID 26578569.
- ^ a b Lujan SA, Williams JS, Kunkel TA (2016-02-01). "Eukaryotic genome instability in light of asymmetric DNA replication". Critical Reviews in Biochemistry and Molecular Biology. 51 (1): 43–52. doi:10.3109/10409238.2015.1117055. PMID 26822554.
- ^ Doublié S, Zahn KE (2014-01-01). "Structural insights into eukaryotic DNA replication". Frontiers in Microbiology. 5: 444. doi:10.3389/fmicb.2014.00444. PMC 4142720. PMID 25202305.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ "NCBI CDD Conserved Protein Domain DNA_polB_delta_exo". www.ncbi.nlm.nih.gov. Retrieved 2016-04-25.
- ^ Brown JA, Suo Z (February 2011). "Unlocking the sugar "steric gate" of DNA polymerases". Biochemistry. 50 (7): 1135–42. doi:10.1021/bi101915z. PMC 3040255. PMID 21226515.
- ^ Krucher NA, Zygmunt A, Mazloum N, Tamrakar S, Ludlow JW, Lee MY (November 2000). "Interaction of the retinoblastoma protein (pRb) with the catalytic subunit of DNA polymerase delta (p125)". Oncogene. 19 (48): 5464–70. doi:10.1038/sj.onc.1203930. PMID 11114723.
- ^ Hogg M, Aller P, Konigsberg W, Wallace SS, Doublié S (January 2007). "Structural and biochemical investigation of the role in proofreading of a beta hairpin loop found in the exonuclease domain of a replicative DNA polymerase of the B family". The Journal of Biological Chemistry. 282 (2): 1432–44. doi:10.1074/jbc.M605675200. PMID 17098747.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Lam YW, Trinkle-Mulcahy L (2015-01-01). "New insights into nucleolar structure and function". F1000prime Reports. 7: 48. doi:10.12703/P7-48. PMC 4447046. PMID 26097721.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Mekhail K, Rivero-Lopez L, Al-Masri A, Brandon C, Khacho M, Lee S (October 2007). "Identification of a common subnuclear localization signal". Molecular Biology of the Cell. 18 (10): 3966–77. doi:10.1091/mbc.E07-03-0295. PMC 1995723. PMID 17652456.
- ^ a b Audas TE, Jacob MD, Lee S (January 2012). "Immobilization of proteins in the nucleolus by ribosomal intergenic spacer noncoding RNA". Molecular Cell. 45 (2): 147–57. doi:10.1016/j.molcel.2011.12.012. PMID 22284675.
- ^ Netz DJ, Stith CM, Stümpfig M, Köpf G, Vogel D, Genau HM, Stodola JL, Lill R, Burgers PM, Pierik AJ (January 2012). "Eukaryotic DNA polymerases require an iron-sulfur cluster for the formation of active complexes". Nature Chemical Biology. 8 (1): 125–32. doi:10.1038/nchembio.721. PMC 3241888. PMID 22119860.
- ^ Paul VD, Lill R (June 2015). "Biogenesis of cytosolic and nuclear iron-sulfur proteins and their role in genome stability". Biochimica et Biophysica Acta. 1853 (6): 1528–39. doi:10.1016/j.bbamcr.2014.12.018. PMID 25583461.
- ^ Gari K, León Ortiz AM, Borel V, Flynn H, Skehel JM, Boulton SJ (July 2012). "MMS19 links cytoplasmic iron-sulfur cluster assembly to DNA metabolism". Science. 337 (6091): 243–5. doi:10.1126/science.1219664. PMID 22678361.
- ^ Stehling O, Vashisht AA, Mascarenhas J, Jonsson ZO, Sharma T, Netz DJ, Pierik AJ, Wohlschlegel JA, Lill R (July 2012). "MMS19 assembles iron-sulfur proteins required for DNA metabolism and genomic integrity". Science. 337 (6091): 195–9. doi:10.1126/science.1219723. PMC 3420340. PMID 22678362.
- ^ Hansen MF, Johansen J, Bjørnevoll I, Sylvander AE, Steinsbekk KS, Sætrom P, Sandvik AK, Drabløs F, Sjursen W (September 2015). "A novel POLE mutation associated with cancers of colon, pancreas, ovaries and small intestine". Familial Cancer. 14 (3): 437–48. doi:10.1007/s10689-015-9803-2. PMC 4559173. PMID 25860647.
- ^ Zuo Y, Deutscher MP (March 2001). "Exoribonuclease superfamilies: structural analysis and phylogenetic distribution". Nucleic Acids Research. 29 (5): 1017–26. doi:10.1093/nar/29.5.1017. PMC 56904. PMID 11222749.
- ^ Simon M, Giot L, Faye G (August 1991). "The 3' to 5' exonuclease activity located in the DNA polymerase delta subunit of Saccharomyces cerevisiae is required for accurate replication". The EMBO Journal. 10 (8): 2165–70. PMC 452904. PMID 1648480.
- ^ Makarova KS, Krupovic M, Koonin EV (2014-01-01). "Evolution of replicative DNA polymerases in archaea and their contributions to the eukaryotic replication machinery". Frontiers in Microbiology. 5: 354. doi:10.3389/fmicb.2014.00354. PMC 4104785. PMID 25101062.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Xie B, Mazloum N, Liu L, Rahmeh A, Li H, Lee MY (November 2002). "Reconstitution and characterization of the human DNA polymerase delta four-subunit holoenzyme". Biochemistry. 41 (44): 13133–42. doi:10.1021/bi0262707. PMID 12403614.
- ^ Lab, Mike Tyers. "Database of Protein, Chemical, and Genetic Interactions | BioGRID". thebiogrid.org. Retrieved 2016-04-25.
- ^ "POLD1 protein (Homo sapiens) - STRING network view". string-db.org. Retrieved 2016-04-25.
- ^ "Cancer Cell Metabolism Database ~~ Bioinformatics and Systems Medicine Laboratory ~~". bioinfo.mc.vanderbilt.edu. Retrieved 2016-04-25.
- ^ "STRING: functional protein association networks". string-db.org. Retrieved 2016-04-25.
- ^ Ono, Keiichiro. "Cytoscape: An Open Source Platform for Complex Network Analysis and Visualization". www.cytoscape.org. Retrieved 2016-04-25.
- ^ "POLD1 protein (Homo sapiens) - STRING network view". string-db.org. Retrieved 2016-04-25.
- ^ a b c Weedon MN, Ellard S, Prindle MJ, Caswell R, Lango Allen H, Oram R, Godbole K, Yajnik CS, Sbraccia P, Novelli G, Turnpenny P, McCann E, Goh KJ, Wang Y, Fulford J, McCulloch LJ, Savage DB, O'Rahilly S, Kos K, Loeb LA, Semple RK, Hattersley AT (August 2013). "An in-frame deletion at the polymerase active site of POLD1 causes a multisystem disorder with lipodystrophy". Nature Genetics. 45 (8): 947–50. doi:10.1038/ng.2670. PMC 3785143. PMID 23770608.
- ^ "Genatlas sheet". genatlas.medecine.univ-paris5.fr. Retrieved 2016-04-25.
- ^ Takahashi Y, Moriwaki S, Sugiyama Y, Endo Y, Yamazaki K, Mori T, Takigawa M, Inoue S (February 2005). "Decreased gene expression responsible for post-ultraviolet DNA repair synthesis in aging: a possible mechanism of age-related reduction in DNA repair capacity". The Journal of Investigative Dermatology. 124 (2): 435–42. doi:10.1111/j.0022-202X.2004.23591.x. PMID 15675965.
- ^ a b Wang JL, Guo HL, Wang PC, Liu CG (December 2012). "Age-dependent down-regulation of DNA polymerase δ1 in human lymphocytes". Molecular and Cellular Biochemistry. 371 (1–2): 157–63. doi:10.1007/s11010-012-1432-6. PMID 22915169.
- ^ Karkhanis V, Wang L, Tae S, Hu YJ, Imbalzano AN, Sif S (August 2012). "Protein arginine methyltransferase 7 regulates cellular response to DNA damage by methylating promoter histones H2A and H4 of the polymerase δ catalytic subunit gene, POLD1". The Journal of Biological Chemistry. 287 (35): 29801–14. doi:10.1074/jbc.M112.378281. PMC 3436169. PMID 22761421.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Cui M, You L, Ren X, Zhao W, Liao Q, Zhao Y (February 2016). "Long non-coding RNA PVT1 and cancer". Biochemical and Biophysical Research Communications. 471 (1): 10–4. doi:10.1016/j.bbrc.2015.12.101. PMID 26850852.
- ^ Li H, Xie B, Zhou Y, Rahmeh A, Trusa S, Zhang S, Gao Y, Lee EY, Lee MY (May 2006). "Functional roles of p12, the fourth subunit of human DNA polymerase delta". The Journal of Biological Chemistry. 281 (21): 14748–55. doi:10.1074/jbc.M600322200. PMID 16510448.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Podust VN, Chang LS, Ott R, Dianov GL, Fanning E (February 2002). "Reconstitution of human DNA polymerase delta using recombinant baculoviruses: the p12 subunit potentiates DNA polymerizing activity of the four-subunit enzyme". The Journal of Biological Chemistry. 277 (6): 3894–901. doi:10.1074/jbc.M109684200. PMID 11711545.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Zhang S, Zhou Y, Trusa S, Meng X, Lee EY, Lee MY (May 2007). "A novel DNA damage response: rapid degradation of the p12 subunit of dna polymerase delta". The Journal of Biological Chemistry. 282 (21): 15330–40. doi:10.1074/jbc.M610356200. PMID 17317665.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Lee MY, Zhang S, Lin SH, Wang X, Darzynkiewicz Z, Zhang Z, Lee EY (2014-01-01). "The tail that wags the dog: p12, the smallest subunit of DNA polymerase δ, is degraded by ubiquitin ligases in response to DNA damage and during cell cycle progression". Cell Cycle. 13 (1): 23–31. doi:10.4161/cc.27407. PMC 3925730. PMID 24300032.
- ^ Zhang S, Zhao H, Darzynkiewicz Z, Zhou P, Zhang Z, Lee EY, Lee MY (October 2013). "A novel function of CRL4(Cdt2): regulation of the subunit structure of DNA polymerase δ in response to DNA damage and during the S phase". The Journal of Biological Chemistry. 288 (41): 29550–61. doi:10.1074/jbc.M113.490466. PMC 3795253. PMID 23913683.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Fan X, Zhang Q, You C, Qian Y, Gao J, Liu P, Chen H, Song H, Chen Y, Chen K, Zhou Y (2014-01-01). "Proteolysis of the human DNA polymerase delta smallest subunit p12 by μ-calpain in calcium-triggered apoptotic HeLa cells". PloS One. 9 (4): e93642. doi:10.1371/journal.pone.0093642. PMC 3972206. PMID 24691096.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Zhang Q, Zhang Q, Chen H, Chen Y, Zhou Y (February 2016). "Multiple forms of human DNA polymerase delta sub-assembling in cellular DNA transactions". Current Protein & Peptide Science. PMID 26916162.
- ^ a b Szekely AM, Chen YH, Zhang C, Oshima J, Weissman SM (October 2000). "Werner protein recruits DNA polymerase delta to the nucleolus". Proceedings of the National Academy of Sciences of the United States of America. 97 (21): 11365–70. doi:10.1073/pnas.97.21.11365. PMC 17206. PMID 11027336.
- ^ a b Karmakar P, Bohr VA (November 2005). "Cellular dynamics and modulation of WRN protein is DNA damage specific". Mechanisms of Ageing and Development. 126 (11): 1146–58. doi:10.1016/j.mad.2005.06.004. PMID 16087220.
- ^ Lee SY, Lee H, Kim ES, Park S, Lee J, Ahn B (April 2015). "WRN translocation from nucleolus to nucleoplasm is regulated by SIRT1 and required for DNA repair and the development of chemoresistance". Mutation Research. 774: 40–8. doi:10.1016/j.mrfmmm.2015.03.001. PMID 25801465.
- ^ Wang X, Zhang S, Zheng R, Yue F, Lin SH, Rahmeh AA, Lee EY, Zhang Z, Lee MY (February 2016). "PDIP46 (DNA polymerase δ interacting protein 46) is an activating factor for human DNA polymerase δ". Oncotarget. 7 (5): 6294–313. doi:10.18632/oncotarget.7034. PMID 26819372.
- ^ Boyer AS, Walter D, Sørensen CS (January 2016). "DNA replication and cancer: From dysfunctional replication origin activities to therapeutic opportunities". Seminars in Cancer Biology. doi:10.1016/j.semcancer.2016.01.001. PMID 26805514.
- ^ Sincennes MC, Humbert M, Grondin B, Lisi V, Veiga DF, Haman A, Cazaux C, Mashtalir N, Affar el B, Verreault A, Hoang T (February 2016). "The LMO2 oncogene regulates DNA replication in hematopoietic cells". Proceedings of the National Academy of Sciences of the United States of America. 113 (5): 1393–8. doi:10.1073/pnas.1515071113. PMC 4747768. PMID 26764384.
- ^ Nick McElhinny SA, Gordenin DA, Stith CM, Burgers PM, Kunkel TA (April 2008). "Division of labor at the eukaryotic replication fork". Molecular Cell. 30 (2): 137–44. doi:10.1016/j.molcel.2008.02.022. PMC 2654179. PMID 18439893.
- ^ a b Johnson RE, Klassen R, Prakash L, Prakash S (July 2015). "A Major Role of DNA Polymerase δ in Replication of Both the Leading and Lagging DNA Strands". Molecular Cell. 59 (2): 163–75. doi:10.1016/j.molcel.2015.05.038. PMC 4517859. PMID 26145172.
- ^ Daigaku Y, Keszthelyi A, Müller CA, Miyabe I, Brooks T, Retkute R, Hubank M, Nieduszynski CA, Carr AM (March 2015). "A global profile of replicative polymerase usage". Nature Structural & Molecular Biology. 22 (3): 192–8. doi:10.1038/nsmb.2962. PMC 4789492. PMID 25664722.
- ^ Pavlov YI, Shcherbakova PV (March 2010). "DNA polymerases at the eukaryotic fork-20 years later". Mutation Research. 685 (1–2): 45–53. doi:10.1016/j.mrfmmm.2009.08.002. PMC 2822129. PMID 19682465.
- ^ Stillman B (July 2015). "Reconsidering DNA Polymerases at the Replication Fork in Eukaryotes". Molecular Cell. 59 (2): 139–41. doi:10.1016/j.molcel.2015.07.004. PMC 4636199. PMID 26186286.
- ^ Burgers PM, Gordenin D, Kunkel TA (February 2016). "Who Is Leading the Replication Fork, Pol ε or Pol δ?". Molecular Cell. 61 (4): 492–3. doi:10.1016/j.molcel.2016.01.017. PMC 4838066. PMID 26895421.
- ^ Korona DA, Lecompte KG, Pursell ZF (March 2011). "The high fidelity and unique error signature of human DNA polymerase epsilon". Nucleic Acids Research. 39 (5): 1763–73. doi:10.1093/nar/gkq1034. PMC 3061053. PMID 21036870.
- ^ Skoneczna A, Kaniak A, Skoneczny M (November 2015). "Genetic instability in budding and fission yeast-sources and mechanisms". FEMS Microbiology Reviews. 39 (6): 917–67. doi:10.1093/femsre/fuv028. PMC 4608483. PMID 26109598.
- ^ Hoopes JI, Cortez LM, Mertz TM, Malc EP, Mieczkowski PA, Roberts SA (February 2016). "APOBEC3A and APOBEC3B Preferentially Deaminate the Lagging Strand Template during DNA Replication". Cell Reports. 14 (6): 1273–82. doi:10.1016/j.celrep.2016.01.021. PMC 4758883. PMID 26832400.
- ^ Reijns MA, Kemp H, Ding J, de Procé SM, Jackson AP, Taylor MS (February 2015). "Lagging-strand replication shapes the mutational landscape of the genome". Nature. 518 (7540): 502–6. doi:10.1038/nature14183. PMC 4374164. PMID 25624100.
- ^ Flood CL, Rodriguez GP, Bao G, Shockley AH, Kow YW, Crouse GF (March 2015). "Replicative DNA polymerase δ but not ε proofreads errors in Cis and in Trans". PLoS Genetics. 11 (3): e1005049. doi:10.1371/journal.pgen.1005049. PMC 4351087. PMID 25742645.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Herr AJ, Kennedy SR, Knowels GM, Schultz EM, Preston BD (March 2014). "DNA replication error-induced extinction of diploid yeast". Genetics. 196 (3): 677–91. doi:10.1534/genetics.113.160960. PMC 3948800. PMID 24388879.
- ^ Morrison A, Johnson AL, Johnston LH, Sugino A (April 1993). "Pathway correcting DNA replication errors in Saccharomyces cerevisiae". The EMBO Journal. 12 (4): 1467–73. PMC 413358. PMID 8385605.
- ^ Li L, Murphy KM, Kanevets U, Reha-Krantz LJ (June 2005). "Sensitivity to phosphonoacetic acid: a new phenotype to probe DNA polymerase delta in Saccharomyces cerevisiae". Genetics. 170 (2): 569–80. doi:10.1534/genetics.104.040295. PMC 1450396. PMID 15802517.
- ^ Meng X, Zhou Y, Zhang S, Lee EY, Frick DN, Lee MY (February 2009). "DNA damage alters DNA polymerase delta to a form that exhibits increased discrimination against modified template bases and mismatched primers". Nucleic Acids Research. 37 (2): 647–57. doi:10.1093/nar/gkn1000. PMC 2632934. PMID 19074196.
- ^ Baranovskiy AG, Lada AG, Siebler HM, Zhang Y, Pavlov YI, Tahirov TH (May 2012). "DNA polymerase δ and ζ switch by sharing accessory subunits of DNA polymerase δ". The Journal of Biological Chemistry. 287 (21): 17281–7. doi:10.1074/jbc.M112.351122. PMC 3366816. PMID 22465957.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Markkanen E, Castrec B, Villani G, Hübscher U (December 2012). "A switch between DNA polymerases δ and λ promotes error-free bypass of 8-oxo-G lesions". Proceedings of the National Academy of Sciences of the United States of America. 109 (50): 20401–6. doi:10.1073/pnas.1211532109. PMC 3528542. PMID 23175785.
- ^ Song J, Hong P, Liu C, Zhang Y, Wang J, Wang P (2015-01-01). "Human POLD1 modulates cell cycle progression and DNA damage repair". BMC Biochemistry. 16: 14. doi:10.1186/s12858-015-0044-7. PMC 4471906. PMID 26087769.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Hocke S, Guo Y, Job A, Orth M, Ziesch A, Lauber K, De Toni EN, Gress TM, Herbst A, Göke B, Gallmeier E (February 2016). "A synthetic lethal screen identifies ATR-inhibition as a novel therapeutic approach for POLD1-deficient cancers". Oncotarget. 7 (6): 7080–95. doi:10.18632/oncotarget.6857. PMID 26755646.
- ^ Miyabe I, Mizuno K, Keszthelyi A, Daigaku Y, Skouteri M, Mohebi S, Kunkel TA, Murray JM, Carr AM (November 2015). "Polymerase δ replicates both strands after homologous recombination-dependent fork restart". Nature Structural & Molecular Biology. 22 (11): 932–8. doi:10.1038/nsmb.3100. PMC 4655445. PMID 26436826.
- ^ Kamath-Loeb AS, Shen JC, Schmitt MW, Loeb LA (April 2012). "The Werner syndrome exonuclease facilitates DNA degradation and high fidelity DNA polymerization by human DNA polymerase δ". The Journal of Biological Chemistry. 287 (15): 12480–90. doi:10.1074/jbc.M111.332577. PMC 3320997. PMID 22351772.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Jansen AM, van Wezel T, van den Akker BE, Ventayol Garcia M, Ruano D, Tops CM, Wagner A, Letteboer TG, Gómez-García EB, Devilee P, Wijnen JT, Hes FJ, Morreau H (December 2015). "Combined mismatch repair and POLE/POLD1 defects explain unresolved suspected Lynch syndrome cancers". European Journal of Human Genetics. doi:10.1038/ejhg.2015.252. PMID 26648449.
- ^ a b c d e f Palles C, Cazier JB, Howarth KM, Domingo E, Jones AM, Broderick P, et al. (February 2013). "Germline mutations affecting the proofreading domains of POLE and POLD1 predispose to colorectal adenomas and carcinomas". Nature Genetics. 45 (2): 136–44. doi:10.1038/ng.2503. PMC 3785128. PMID 23263490.
- ^ a b c d Valle L, Hernández-Illán E, Bellido F, Aiza G, Castillejo A, Castillejo MI, et al. (July 2014). "New insights into POLE and POLD1 germline mutations in familial colorectal cancer and polyposis". Human Molecular Genetics. 23 (13): 3506–12. doi:10.1093/hmg/ddu058. PMID 24501277.
- ^ a b c d Bellido F, Pineda M, Aiza G, Valdés-Mas R, Navarro M, Puente DA, Pons T, González S, Iglesias S, Darder E, Piñol V, Soto JL, Valencia A, Blanco I, Urioste M, Brunet J, Lázaro C, Capellá G, Puente XS, Valle L (April 2016). "POLE and POLD1 mutations in 529 kindred with familial colorectal cancer and/or polyposis: review of reported cases and recommendations for genetic testing and surveillance". Genetics in Medicine. 18 (4): 325–32. doi:10.1038/gim.2015.75. PMC 4823640. PMID 26133394.
- ^ a b c d Briggs S, Tomlinson I (June 2013). "Germline and somatic polymerase ε and δ mutations define a new class of hypermutated colorectal and endometrial cancers". The Journal of Pathology. 230 (2): 148–53. doi:10.1002/path.4185. PMC 3709119. PMID 23447401.
- ^ a b c Church DN, Briggs SE, Palles C, Domingo E, Kearsey SJ, Grimes JM, Gorman M, Martin L, Howarth KM, Hodgson SV, Kaur K, Taylor J, Tomlinson IP (July 2013). "DNA polymerase ε and δ exonuclease domain mutations in endometrial cancer". Human Molecular Genetics. 22 (14): 2820–8. doi:10.1093/hmg/ddt131. PMC 3690967. PMID 23528559.
- ^ a b Heitzer E, Tomlinson I (February 2014). "Replicative DNA polymerase mutations in cancer". Current Opinion in Genetics & Development. 24: 107–13. doi:10.1016/j.gde.2013.12.005. PMC 4003352. PMID 24583393.
- ^ a b Shinbrot E, Henninger EE, Weinhold N, Covington KR, Göksenin AY, Schultz N, Chao H, Doddapaneni H, Muzny DM, Gibbs RA, Sander C, Pursell ZF, Wheeler DA (November 2014). "Exonuclease mutations in DNA polymerase epsilon reveal replication strand specific mutation patterns and human origins of replication". Genome Research. 24 (11): 1740–50. doi:10.1101/gr.174789.114. PMC 4216916. PMID 25228659.
- ^ Arora S, Yan H, Cho I, Fan HY, Luo B, Gai X, Bodian DL, Vockley JG, Zhou Y, Handorf EA, Egleston BL, Andrake MD, Nicolas E, Serebriiskii IG, Yen TJ, Hall MJ, Golemis EA, Enders GH (December 2015). "Genetic Variants That Predispose to DNA Double-Strand Breaks in Lymphocytes From a Subset of Patients With Familial Colorectal Carcinomas". Gastroenterology. 149 (7): 1872–1883.e9. doi:10.1053/j.gastro.2015.08.052. PMC 4663158. PMID 26344056.
- ^ Waterfall JJ, Meltzer PS (March 2015). "Avalanching mutations in biallelic mismatch repair deficiency syndrome". Nature Genetics. 47 (3): 194–6. doi:10.1038/ng.3227. PMID 25711864.
- ^ a b Schlesner M, Eils R (2015-01-01). "Hypermutation takes the driver's seat". Genome Medicine. 7 (1): 31. doi:10.1186/s13073-015-0159-x. PMC 4376156. PMID 25821521.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Shlien A, Campbell BB, de Borja R, Alexandrov LB, Merico D, Wedge D, et al. (March 2015). "Combined hereditary and somatic mutations of replication error repair genes result in rapid onset of ultra-hypermutated cancers". Nature Genetics. 47 (3): 257–62. doi:10.1038/ng.3202. PMID 25642631.
- ^ Bouffet E, Larouche V, Campbell BB, Merico D, de Borja R, Aronson M, et al. (March 2016). "Immune Checkpoint Inhibition for Hypermutant Glioblastoma Multiforme Resulting From Germline Biallelic Mismatch Repair Deficiency". Journal of Clinical Oncology. doi:10.1200/JCO.2016.66.6552. PMID 27001570.
- ^ Howitt BE, Shukla SA, Sholl LM, Ritterhouse LL, Watkins JC, Rodig S, Stover E, Strickland KC, D'Andrea AD, Wu CJ, Matulonis UA, Konstantinopoulos PA (December 2015). "Association of Polymerase e-Mutated and Microsatellite-Instable Endometrial Cancers With Neoantigen Load, Number of Tumor-Infiltrating Lymphocytes, and Expression of PD-1 and PD-L1". JAMA Oncology. 1 (9): 1319–23. doi:10.1001/jamaoncol.2015.2151. PMID 26181000.
- ^ van Gool IC, Eggink FA, Freeman-Mills L, Stelloo E, Marchi E, de Bruyn M, Palles C, Nout RA, de Kroon CD, Osse EM, Klenerman P, Creutzberg CL, Tomlinson IP, Smit VT, Nijman HW, Bosse T, Church DN (July 2015). "POLE Proofreading Mutations Elicit an Antitumor Immune Response in Endometrial Cancer". Clinical Cancer Research. 21 (14): 3347–55. doi:10.1158/1078-0432.CCR-15-0057. PMC 4627582. PMID 25878334.
- ^ Khanna A (June 2015). "DNA damage in cancer therapeutics: a boon or a curse?". Cancer Research. 75 (11): 2133–8. doi:10.1158/0008-5472.CAN-14-3247. PMID 25931285.
- ^ Roberts SA, Gordenin DA (December 2014). "Hypermutation in human cancer genomes: footprints and mechanisms". Nature Reviews. Cancer. 14 (12): 786–800. doi:10.1038/nrc3816. PMC 4280484. PMID 25568919.
- ^ Roos WP, Thomas AD, Kaina B (January 2016). "DNA damage and the balance between survival and death in cancer biology". Nature Reviews. Cancer. 16 (1): 20–33. doi:10.1038/nrc.2015.2. PMID 26678314.
- ^ da Costa LT, Liu B, el-Deiry W, Hamilton SR, Kinzler KW, Vogelstein B, Markowitz S, Willson JK, de la Chapelle A, Downey KM (January 1995). "Polymerase delta variants in RER colorectal tumours". Nature Genetics. 9 (1): 10–1. doi:10.1038/ng0195-10. PMID 7704014.
- ^ Flohr T, Dai JC, Büttner J, Popanda O, Hagmüller E, Thielmann HW (March 1999). "Detection of mutations in the DNA polymerase delta gene of human sporadic colorectal cancers and colon cancer cell lines". International Journal of Cancer. 80 (6): 919–29. doi:10.1002/(sici)1097-0215(19990315)80:6<919::aid-ijc19>3.0.co;2-u. PMID 10074927.
- ^ Preston BD, Albertson TM, Herr AJ (October 2010). "DNA replication fidelity and cancer". Seminars in Cancer Biology. 20 (5): 281–93. doi:10.1016/j.semcancer.2010.10.009. PMC 2993855. PMID 20951805.
- ^ Popanda O, Flohr T, Fox G, Thielmann HW (November 1999). "A mutation detected in DNA polymerase delta cDNA from Novikoff hepatoma cells correlates with abnormal catalytic properties of the enzyme". Journal of Cancer Research and Clinical Oncology. 125 (11): 598–608. doi:10.1007/s004320050322. PMID 10541966.
- ^ Venkatesan RN, Treuting PM, Fuller ED, Goldsby RE, Norwood TH, Gooley TA, Ladiges WC, Preston BD, Loeb LA (November 2007). "Mutation at the polymerase active site of mouse DNA polymerase delta increases genomic instability and accelerates tumorigenesis". Molecular and Cellular Biology. 27 (21): 7669–82. doi:10.1128/MCB.00002-07. PMC 2169052. PMID 17785453.
- ^ a b c Pelosini C, Martinelli S, Ceccarini G, Magno S, Barone I, Basolo A, Fierabracci P, Vitti P, Maffei M, Santini F (November 2014). "Identification of a novel mutation in the polymerase delta 1 (POLD1) gene in a lipodystrophic patient affected by mandibular hypoplasia, deafness, progeroid features (MDPL) syndrome". Metabolism. 63 (11): 1385–9. doi:10.1016/j.metabol.2014.07.010. PMID 25131834.
- ^ Reinier F, Zoledziewska M, Hanna D, Smith JD, Valentini M, Zara I, Berutti R, Sanna S, Oppo M, Cusano R, Satta R, Montesu MA, Jones C, Cerimele D, Nickerson DA, Angius A, Cucca F, Cottoni F, Crisponi L (November 2015). "Mandibular hypoplasia, deafness, progeroid features and lipodystrophy (MDPL) syndrome in the context of inherited lipodystrophies". Metabolism. 64 (11): 1530–40. doi:10.1016/j.metabol.2015.07.022. PMID 26350127.
- ^ Oshima J, Sidorova JM, Monnat RJ (March 2016). "Werner syndrome: Clinical features, pathogenesis and potential therapeutic interventions". Ageing Research Reviews. doi:10.1016/j.arr.2016.03.002. PMID 26993153.
- ^ Lessel D, Hisama FM, Szakszon K, Saha B, Sanjuanelo AB, Salbert BA, Steele PD, Baldwin J, Brown WT, Piussan C, Plauchu H, Szilvássy J, Horkay E, Högel J, Martin GM, Herr AJ, Oshima J, Kubisch C (November 2015). "POLD1 Germline Mutations in Patients Initially Diagnosed with Werner Syndrome". Human Mutation. 36 (11): 1070–9. doi:10.1002/humu.22833. PMC 4684254. PMID 26172944.
- ^ Pachlopnik Schmid J, Lemoine R, Nehme N, Cormier-Daire V, Revy P, Debeurme F, Debré M, Nitschke P, Bole-Feysot C, Legeai-Mallet L, Lim A, de Villartay JP, Picard C, Durandy A, Fischer A, de Saint Basile G (December 2012). "Polymerase ε1 mutation in a human syndrome with facial dysmorphism, immunodeficiency, livedo, and short stature ("FILS syndrome")". The Journal of Experimental Medicine. 209 (13): 2323–30. doi:10.1084/jem.20121303. PMC 3526359. PMID 23230001.
- ^ Thiffault I, Saunders C, Jenkins J, Raje N, Canty K, Sharma M, Grote L, Welsh HI, Farrow E, Twist G, Miller N, Zwick D, Zellmer L, Kingsmore SF, Safina NP (2015-01-01). "A patient with polymerase E1 deficiency (POLE1): clinical features and overlap with DNA breakage/instability syndromes". BMC Medical Genetics. 16: 31. doi:10.1186/s12881-015-0177-y. PMC 4630961. PMID 25948378.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Guénantin AC, Briand N, Bidault G, Afonso P, Béréziat V, Vatier C, Lascols O, Caron-Debarle M, Capeau J, Vigouroux C (May 2014). "Nuclear envelope-related lipodystrophies". Seminars in Cell & Developmental Biology. 29: 148–57. doi:10.1016/j.semcdb.2013.12.015. PMID 24384368.
- ^ a b "GeneTests.org". GeneTests.org. Retrieved 2016-04-25.
- ^ "MDP syndrome caused by a change in the POLD1 gene".