
PIM2 (gene)






Serine/threonine-protein kinase Pim-2 is an enzyme that in humans is encoded by the PIM2 .[5][6]
PIM2 or Proviral Integrations of Moloney virus 2 is serine/threonine kinase that has roles in cell growth, proliferation, apoptosis, and regulation of signal transduction cascades.[7]
Structure[edit]
Thus far, most of the structural information pertaining to the PIM kinase family has been limited to PIM1. As a result, most of inhibitor development efforts has also been towards PIM1. PIM2 shares 55% sequence identity with PIM1, and the structure of PIM2 is quite closely related to PIM1.[8] Like PIM1, PIM2 shows a bi-lobal kinase architecture with a constitutively active closed conformation. The main chain of both molecules is identical with the exception of two flexible regions in the N-terminal lobe.

The most significant structural difference between PIM1 and PIM2 is the absence of the terminal αJ helix in PIM2. The last 23 residues of PIM2 are quite different from PIM1, as PIM2 contains 6 proline residues in this region and is not believed to form the same tertiary structures. As a result, the absence of the interactions present in this region may increase flexibility in PIM2 within the N-terminal kinase lobe and contribute to the disordered regions of the PIM2 structure.[9]
Function[edit]
PIM2 is expressed with high levels in the brain and lymphoid cells. PIM1-3 compound knockout mice that survived the perinatal period showed a large reduction in body size. This suggests that the PIM enzymes are important for body growth.[10] Experiments have implicated that PIM1 and PIM2 are necessary for cytokine-dependent proliferation and survival of lymphocytes.[11] Experiments with transgenic mice with induced lymphomas revealed elevated levels of PIM2 as a frequent but late event in tumorigenesis.[12]
Experiments done on nuclear factor κB (NFκB) nuclear translocation in human perineural invasion (PNI) revealed that an up-regulation of NFκB and its downstream target, PIM2, were components of an antiapoptosis signaling cascade, which is associated with cancer cells in PNI. This cascade may regulate the inhibition of apoptosis. The study also showed that elevated levels of PIM2 have been associated with PNI.[13] The PIM2 kinase has therefore emerged as a key drug target to restore apoptosis in drug resistant human cancers.[14][15][16]
Mechanism[edit]
In reported crystal structures, PIM1 and PIM2 assume an active conformation. Typically, kinases’ active state is characterized by the presence of the conserved lysine, a closed lobe conformation, and a well-structured activation segment. The activation segment often necessitates phosphorylation in order for there to be catalytic activity. Once phosphorylated, the active segment folds onto the lower lobe and reorganizes the peptide-binding site, which consequently leads to enzymatic activation. However, PIM kinases are catalytically active without phosphorylation. The crystal structures show that the unphosphorylated activation segment forms many polar interactions with the lower kinase lobe, which stabilizes the active conformation. While PIM kinase do autophosphorylate, the functional consequences are not known.[17]
Inhibitors[edit]
-
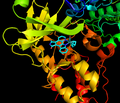 Inhibitor acts as ATP mimetic in ATP binding pocket
Inhibitor acts as ATP mimetic in ATP binding pocket -
 Staurosporine and Ruthenium Compound 12
Staurosporine and Ruthenium Compound 12 -
 General Structure of Organoruthenium inhibitor
General Structure of Organoruthenium inhibitor



PIM2 (along with PIM1) has a unique binding pocket for ATP with a hinged region, making it an attractive target for potent small-molecule PIM kinase inhibitors.[18]
Many inhibitors are often more selective for PIM1 and PIM3 over PIM2. In other words, PIM2 is usually inhibited with much lower potency. Thus far, structural models are unable to explain this phenomenon. However, it could be related to the differences in the dynamic properties of the different PIM isoforms.
In a series of organoruthenium compounds [19] based on a Staurosporine scaffold[20] compound 12 gave almost complete inhibition at a concentration of 10 nM. However, it was marginally less effective against PIM1.
The SAR suggests that the addition of potential hydrogen bonding groups at the R1 and R2 positions dramatically increases potency against both kinases. Similar substitution of the R3 position was less effective and halogen substitution was even more disruptive.
Clinical significance[edit]
In a study with 48 patients who had non-Hodgkin's lymphoma (NHL) and lymphocytic leukemia, hPim-2 expression was analyzed using in-situ hybridization, quantitative RT-PCR and FACS analysis. The studies showed higher levels of expression in NHL over normal lymphocytes as well as in chronic lymphocytic leukemia over normal B-Cells.[21]
Elevated PIM2 levels have also been found in primary blasts from acute myeloid leukemia patients. PIM2 may be an important kinase in the phosphorylation of 4E-BP1. Constitutive phosphorylation of 4E-BP1 is commonly found in cancers and contributes to the sustained translation of malignancy related transcripts, among which are c-Myc and Cyclin D. Knockdown of PIM2 by iRNA strongly reduced the accumulation of oncogenic proteins.[22] As a result, PIM2 may be an attractive target for acute myeloid leukemia.
References[edit]
- ^ a b c GRCh38: Ensembl release 89: ENSG00000102096 – Ensembl, May 2017
- ^ a b c GRCm38: Ensembl release 89: ENSMUSG00000031155 – Ensembl, May 2017
- ^ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ Baytel D, Shalom S, Madgar I, Weissenberg R, Don J (Dec 1998). "The human Pim-2 proto-oncogene and its testicular expression". Biochim Biophys Acta. 1442 (2–3): 274–85. doi:10.1016/s0167-4781(98)00185-7. PMID 9804974.
- ^ "Entrez Gene: PIM2 pim-2 oncogene".
- ^ Alvarado Y, Giles FJ, Swords RT (2012). "The PIM kinases in hematological cancers". Expert Rev Hematol. 5 (1): 81–96. doi:10.1586/ehm.11.69. PMID 22272708. S2CID 29518294.
- ^ Debreczeni JE, Bullock AN, Atilla GE, Williams DS, Bregman H, Knapp S, Meggers E (Feb 2006). "Ruthenium half-sandwich complexes bound to protein kinase Pim-1". Angewandte Chemie. 45 (10): 1580–5. doi:10.1002/anie.200503468. PMID 16381041.
- ^ Bullock AN, Russo S, Amos A, Pagano N, Bregman H, Debreczeni JE, Lee WH, von Delft F, Meggers E, Knapp S (2005). "Crystal Structure of the PIM2 Kinase in Complex with an Organoruthenium Inhibitor". PLOS ONE. 4 (10): e7112. doi:10.1371/journal.pone.0007112. PMC 2743286. PMID 19841674.
- ^ Mikkers H, Nawijn M, Allen J, Brouwers C, Verhoeven E, Jonkers J, Berns A (2004). "Mice Deficient for All PIM Kinases Display Reduced Body Size and Impaired Responses to Hematopoietic Growth Factors". Mol Cell Biol. 24 (13): 6104–6115. doi:10.1128/MCB.24.13.6104-6115.2004. PMC 480904. PMID 15199164.
- ^ Tahvanainen J, Kyläniemi MK, Kanduri K, Gupta B, Lähteenmäki H, Kallonen T, Rajavuori A, Rasool O, Koskinen PJ, Rao KV, Lähdesmäki H, Lahesmaa R (2013). "Proviral Integration Site for Moloney Murine Leukemia Virus (PIM) Kinases Promote Human T Helper 1 Cell Differentiation". The Journal of Biological Chemistry. 288 (5): 3048–3058. doi:10.1074/jbc.M112.361709. PMC 3561529. PMID 23209281.
- ^ Breuer ML, Cuypers HT, Berns A (Mar 1989). "Evidence for the involvement of pim-2, a new common proviral insertion site, in progression of lymphomas". The EMBO Journal. 8 (3): 743–8. doi:10.1002/j.1460-2075.1989.tb03434.x. PMC 400870. PMID 2721500.
- ^ Ayala GE, Dai H, Ittmann M, Li R, Powell M, Frolov A, et al. (September 2004). "Growth and survival mechanisms associated with perineural invasion in prostate cancer". Cancer Research. 64 (17): 6082–90. doi:10.1158/0008-5472.CAN-04-0838. PMID 15342391.
- ^ Giles F (2005). "A Pim kinase inhibitor, please". Blood. 105 (11): 4158–4159. doi:10.1182/blood-2005-03-1150.
- ^ Malone T, Schäfer L, Simon N, Heavey S, Cuffe S, Finn S, et al. (March 2020). "Current perspectives on targeting PIM kinases to overcome mechanisms of drug resistance and immune evasion in cancer" (PDF). Pharmacology & Therapeutics. 207: 107454. doi:10.1016/j.pharmthera.2019.107454. PMID 31836451. S2CID 209357486.
- ^ Luszczak S, Kumar C, Sathyadevan VK, Simpson BS, Gately KA, Whitaker HC, Heavey S (2020). "PIM kinase inhibition: co-targeted therapeutic approaches in prostate cancer". Signal Transduction and Targeted Therapy. 5: 7. doi:10.1038/s41392-020-0109-y. PMC 6992635. PMID 32025342.
- ^ Brault L, Gasser C, Bracher F, Huber K, Knapp S, Schwaller J (2010). "PIM serine/threonine kinases in the pathogenesis and therapy of hematologic malignancies and solid cancers". Haematologica. 95 (6): 1004–1015. doi:10.3324/haematol.2009.017079. PMC 2878801. PMID 20145274.
- ^ Pogacic V, Bullock AN, Fedorov O, Filippakopoulos P, Gasser C, Biondi A, Meyer-Monard S, Knapp S, Schwaller J (Jul 2007). "Structural analysis identifies imidazo[1,2-b]pyridazines as PIM kinase inhibitors with in vitro antileukemic activity". Cancer Research. 67 (14): 6916–24. doi:10.1158/0008-5472.CAN-07-0320. PMID 17638903.
- ^ Pagano N, Maksimoska J, Bregman H, Williams DS, Webster RD, Xue F, Meggers E (Apr 2007). "Ruthenium half-sandwich complexes as protein kinase inhibitors: derivatization of the pyridocarbazole pharmacophore ligand". Organic & Biomolecular Chemistry. 5 (8): 1218–27. doi:10.1039/b700433h. PMID 17406720.
- ^ Meggers E (Jun 2007). "Exploring biologically relevant chemical space with metal complexes". Current Opinion in Chemical Biology. 11 (3): 287–92. doi:10.1016/j.cbpa.2007.05.013. PMID 17548234.
- ^ Cohen AM, Grinblat B, Bessler H, Kristt D, Kremer A, Schwartz A, Halperin M, Shalom S, Merkel D, Don J (2004). "Increased Expression of the hPim-2 Gene In Human Chronic lymphocytic Leukemia and Non-Hodgkin Lymphoma". Leukemia & Lymphoma. 45 (5): 951–955. doi:10.1080/10428190310001641251. PMID 15291354. S2CID 24369286.
- ^ Moerke NJ, Aktas H, Chen H, Cantel S, Reibarkh MY, Fahmy A, Gross JD, Degterev A, Yuan J, Chorev M, Halperin JA, Wagner G (Jan 2007). "Small-molecule inhibition of the interaction between the translation initiation factors eIF4E and eIF4G". Cell. 128 (2): 257–67. doi:10.1016/j.cell.2006.11.046. PMID 17254965.
Further reading[edit]
- Yan B, Zemskova M, Holder S, Chin V, Kraft A, Koskinen PJ, Lilly M (2003). "The PIM-2 kinase phosphorylates BAD on serine 112 and reverses BAD-induced cell death". J. Biol. Chem. 278 (46): 45358–67. doi:10.1074/jbc.M307933200. PMID 12954615.
- Cohen AM, Grinblat B, Bessler H, Kristt D, Kremer A, Schwartz A, Halperin M, Shalom S, Merkel D, Don J (2004). "Increased expression of the hPim-2 gene in human chronic lymphocytic leukemia and non-Hodgkin lymphoma". Leuk. Lymphoma. 45 (5): 951–5. doi:10.1080/10428190310001641251. PMID 15291354. S2CID 24369286.
- Hammerman PS, Fox CJ, Cinalli RM, Xu A, Wagner JD, Lindsten T, Thompson CB (2005). "Lymphocyte transformation by Pim-2 is dependent on nuclear factor-kappaB activation". Cancer Res. 64 (22): 8341–8. doi:10.1158/0008-5472.CAN-04-2284. PMID 15548703.
- Rainio EM, Ahlfors H, Carter KL, Ruuska M, Matikainen S, Kieff E, Koskinen PJ (2005). "Pim kinases are upregulated during Epstein-Barr virus infection and enhance EBNA2 activity". Virology. 333 (2): 201–6. doi:10.1016/j.virol.2005.01.001. PMID 15721354.
- Dai H, Li R, Wheeler T, Diaz de Vivar A, Frolov A, Tahir S, Agoulnik I, Thompson T, Rowley D, Ayala G (2005). "Pim-2 upregulation: biological implications associated with disease progression and perinueral invasion in prostate cancer". Prostate. 65 (3): 276–86. doi:10.1002/pros.20294. PMID 16015593. S2CID 44856689.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. Bibcode:2005Natur.437.1173R. doi:10.1038/nature04209. PMID 16189514. S2CID 4427026.
- Macdonald A, Campbell DG, Toth R, McLauchlan H, Hastie CJ, Arthur JS (2006). "Pim kinases phosphorylate multiple sites on Bad and promote 14-3-3 binding and dissociation from Bcl-XL". BMC Cell Biol. 7: 1. doi:10.1186/1471-2121-7-1. PMC 1368972. PMID 16403219.
External links[edit]
- Overview of all the structural information available in the PDB for UniProt: Q9P1W9 (Serine/threonine-protein kinase pim-2) at the PDBe-KB.
PDB gallery | |
|---|---|
|
